

Volume 11, Issue 3 (2025)
Pharm Biomed Res 2025, 11(3): 215-226 |
Back to browse issues page



Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Mozirandi W, Fru G C, Mukanganyama S. Inhibition of Biofilms by Extracts From Vernonia adoensis in Pseudomonas aeruginosa and Staphylococcus aureus. Pharm Biomed Res 2025; 11 (3) :215-226
URL: http://pbr.mazums.ac.ir/article-1-551-en.html
URL: http://pbr.mazums.ac.ir/article-1-551-en.html



1- Department of Biotechnology and Biochemistry, University of Zimbabwe, Harare, Zimbabwe.
2- Department of Chemistry, University of Buea, Buea, Cameroon.
2- Department of Chemistry, University of Buea, Buea, Cameroon.
Full-Text [PDF 895 kb]
(213 Downloads)
| Abstract (HTML) (1054 Views)
Discussion
This study aimed to investigate the effects of V. adoensis leaf extracts on S. aureus and P. aeruginosa biofilms and determine the phytochemical composition of the extract by UPLC-MS. The leaf extract displayed antibiofilm potential against formation of biofilms by both P. aeruginosa and S. aureus. Since bacterial resistance to antibiotics and their survival are associated with their ability to form biofilms [32], extracts that have the potential to decrease biofilm formation would be useful and can be used in conjunction with the antibiotics to decrease bacterial resistance. The results of this study are similar to other studies which have also been done and shown that extracts are capable of inhibiting formation of biofilms by bacteria [33, 14].
When biofilm-forming pathogens such as P. aeruginosa and S. aureus form biofilms in patients, despite the aggressive use of antibiotics, colonization is often a life-long problem leading to chronic diseases [7]. Therefore, preventing the pathogen’s transition from the planktonic state to the biofilm growth mode is the most important step for combating biofilm-associated pathogens, as the ability of pathogens to resist antibiotics is significantly enhanced 10 to 1000 times once they form biofilms [32]
Clinically, biofilm inhibitors can be used directly to reduce virulence factors from the infectious bacteria [3] or to treat biofilm along with conventional antibiotic [33]. The extract used in this study has shown the ability to destruct mature biofilms of P. aeruginosa; therefore, it may be useful in destruction or reduction of preformed biofilms of the bacteria. The weakening or destruction of the mature biomass can be followed by washout or subsequent biocidal inactivation of the detached biomass [34]. Weakening or destroying the mature biofilm will render cells susceptible to antibiotics. It was observed that all the tested concentrations of the extract significantly inhibited metabolic activity on bacteria in preformed biofilms for all tested strains. Plants contain various secondary metabolites, some of which have been found to exert a range of biological activities [35, 36]. The antibiofilm activity produced by the extracts could be due to the presence of individual phytochemicals that are active against S. aureus and P. aeruginosa biofilm or could be the result of the synergistic effect of two or more phytochemicals contained in the extract.
Analysis of plant extracts helps to determine whether the plant species contains any individual compound or group of compounds, which may substantiate its current commercial and traditional use as herbal medicine. Further phytochemical analysis also helps to determine the most appropriate methods of extracting these compounds. Chromatographic techniques contribute significantly to natural products, especially regarding identification, separation, and characterization of bioactive compounds from plant sources [37]. In this study, UPLC-MS was used and UPLC- MS can identify semipolar metabolites with advantages of high precision and short time consumed.
In this study, UPLC-MS analysis of crude acetone extract of V. adoensis revealed the presence of more than five phytochemicals, most of which had been shown to possess some pharmacological properties. For instance, kaempferol, one of the compounds identified in the study, has been shown to exhibit good antioxidant activity [38, 39], anti-inflammatory [40] and anticancer activity [41]. Kaempferol had also been previously identified from Arum palaestinum (Araceae) leaves in a similar study by Abu-reidar [42]. Another compound identified by UPLC-MS was quinic acid which has been shown to have potent anti-herpes activity [43]. Quinic acid has also been previously identified in a similar study [42]. UPLC-MS also identified the presence of caffeic acid in V. adoensis leaves and this phytocompound has been reported to be a potential anti-inflammatory and anticancer agent [44, 45]. A similar study identified caffeic acid in plant species (Hyptis suaveolens and Boerhavia diffusa leaves) [29]. Some compounds identified in the extract by UPLC-MS including rhamnetin and luteon have also been shown to possess biological activity. Rhamnetin was shown to have anti-inflammatory as well as free radical scavenging activity [46, 47]. Luteon has been shown to possess anti-inflammatory [48] and anticancer activities [49]. Similar studies had also identified rhamnetin in betula [50] while Luteolin was identified from burdock, Arctium lappa L leaves [51]. The phytochemical profiles may be used as a pharmacognostical tool for the identification of the plants and as a starting point in the construction of chromatographic fingerprints which can be useful for evaluating the quality of herbal medicines [52, 53]. The observed antibiofilm activity of the extract can be attributed to the different phytochemical constituents that have been found to present in it.
Conclusion
Leaf extracts from V. adoensis have antibiofilm activity on P. aeruginosa and S. aureus biofilms. The phytochemicals identified in V. adoensis extracts may play major roles in the reported biological activities and pharmacological properties of the plant and could lead to novel drugs with potent antibiofilm activity. Further follow-up studies are required in the future in order to elucidate the underlying mechanism of the antibiofilm activity of the extract.
Ethical Considerations
Compliance with ethical guidelines
The study was conducted according to the protocol approved by the Faculty of Science Higher Degrees Committee University of Zimbabwe, Harare, Zimbabwe (Code: Paper HD/166 of 2016).
Funding
This study was supported by the Swedish International Development Agency via the International Science Programs (ISP), and through the International Program in the Chemical Sciences Uppsala University, Uppsala, Sweden (Code: ISP IPICS: ZIM01) and the International Foundation for Sciences (IFS), Stockholm, Sweden (Code: IFS F/3413-03F). This study was supported by the Alliance for Global Health and Science (University of California, Berkeley, Unites States).
Authors' contributions
Conceptualization, study design, and project administration: Stanley Mukanganyama; Experiments: Winnie Mozirandi; Data interpretation: Chi Godloves Fru; Writing and final approval: All authors.
Conflict of interest
The authors declared no conflict of interest.
Acknowledgments
The authors acknowledge the assistance of Christopher Chapano, a taxonomist at the National Herbarium and Botanical Gardens, Harare, Zimbabwe, in authenticating the plant sample names.
Full-Text: (223 Views)
Introduction
An increase in microbial infections has increased morbidity and mortality in healthcare settings [1]. The rise of antibiotic resistance in bacteria is reported to cause an increase in nosocomial infections each year [2]. Biofilms are estimated to be responsible for more than 65% of nosocomial infections and 80% of all microbial infections [3, 4]. A biofilm is an assemblage of surface-associated microbial cells that are enclosed in an extracellular polymeric substance matrix [5]. Biofilm-related infections are difficult to eradicate as there is an increase in antimicrobial tolerance in biofilms compared to planktonic cells [6, 7]. The ability of cells in a biofilm to exert higher resistance to antimicrobials can be attributed to inactivation and reduction of penetration of active antibiotic compounds into the inner layers of the biofilms and persister cells. The increased resistance of cells in biofilm form can also be attributed to the slow growth rate of cells in biofilms and their altered metabolism, oxygen gradients, and extracellular biofilm matrix [4]. Pseudomonas aeruginosa and Staphylococcus aureus are among the most common pathogens found in biofilms in a healthcare setting [7]. These two bacterial species have been significantly identified in nosocomial infections, which have become more rampant and deadly [8]. P. aeruginosa and S. aureus are characterized by potential drug resistance mechanisms, and of major concern is their ability to form biofilms [9]. Antibiotic treatment of biofilm-related infections often provided relief from the symptoms of the infection but may not necessarily cure the infection, mainly due to the biofilms that act as reservoirs for disease recurrence [10]. Biofilms can have important negative effects on human health, for example, P. aeruginosa forms biofilms in the lungs of patients with cystic fibrosis, leading to chronic inflammation and lung tissue damage [7]. The biofilms of P. aeruginosa also cause the persistence of chronic wound infections [11]. Biofilms of S. aureus are responsible for S. aureus-related implant infections [12].
Although biofilms often resist to conventional antibiotic therapy, there are promising alternatives considering the reported efficacy of various plant metabolites against bacteria in biofilms. Phytochemicals from medicinal plants that show antimicrobial activities have the potential to be alternative antimicrobial agents, as their structures differ from those of the more studied microbial sources [13, 14]. Therefore, their mode of action may very likely differ [15]. Vernonia adoensis is a commonly used African ethnomedicinal plant that has been proven to have antimicrobial activity [16, 17]. Screening active compounds from some Vernonia species has led to the discovery of new lead compounds which have efficient protection and treatment roles, including anticancer [18], antiplasmodial [19] and antibacterial [20] activity. Primary phytochemical screening of V. adoensis collected from Kenya has shown some important pharmacological phytochemicals, such as phenols, saponnins, flavonoids, glycosides, tannins, alkaloids, and terpenoids [21]. The composition of phytochemical components varies with geographical location and harvesting conditions [22, 23]. Therefore, the current study aimed to determine the possible chemical components of phytoconstituents from V. adoensis leaves collected from Zimbabwe using gas chromatography–mass spectrometry (GC-MS) and ultra-performance liquid chromatography-mass spectroscopy (UPLC-MS) techniques, and to evaluate the antibiofilm activity of the phytochemicals.
Materials and Methods
Plant material collection
V. adoensis was collected in Centenary, (Geographic coordinates, Latitude: 16º43’22’’ S, Longitude: 31º06’52’’ E, elevation above sea level: 1156 m) Mashonaland Central Province of Zimbabwe. Plant collection was performed in March 2018. Authentication and classification of the plant were performed by a taxonomist from the National Herbarium and Botanic Garden (Harare, Zimbabwe). Herbarium samples C1 E7, were kept at the National Herbarium and Botanic Garden (Harare, Zimbabwe) and the Department of Biochemistry, University of Zimbabwe.
Preparation of acetone extract
The leaf acetone extract of V. adoensis has been reported to have an inhibitory effect on the growth of both P. aeruginosa and S. aureus [17]; therefore, the acetone extract was used to evaluate the effect of V. adoensis on biofilms of S. aureus and P. aeruginosa. The extract was prepared using Mozirandi et al. 2019 [20].
V. adoensis leaves were washed under running tap water and dried in the oven at 40 oC. The dried leaves were pounded in a clean mortar and further ground in a two-speed blender (Cole Parmer Instrument CO., Vernon Hills, USA) to obtain a fine powder to which 10 mL of acetone (Sigma-Aldrich, Taufkirchen, Germany) was added to each 1 g of plant material to extract phytochemicals by maceration [24]. The extract was then filtered using Whatman’s no one filter paper (Sigma-Aldrich, Taufkirchen, Germany). The filtrate was concentrated to dryness by evaporation at room temperature in a fume hood with air stream. The dry extract was stored at 4 ºC for further use.
Bacteria and culture conditions
S. aureus (ATCC 9144) and P. aeruginosa (ATCC 27853), obtained from the Division of Microbiology, Department of Biological Sciences, University of Botswana were separately inoculated in TSB supplemented with 1% glucose and cultured overnight at 37 ºC in a shaking incubator at 120 rpm (SI300 Incubated shaker, Jeiotech, Korea). Cells were centrifuged at 3500 rpm for 4 min on a Hettich Rotofix 32 centrifuge (Tuttlingen, Germany). The pellet was then suspended in fresh media and a 0.5 McFarland Standard was used to create inoculum densities of 5×108 CFU/mL.
Effect of the extract on biofilm formation
Evaluation of the effect of the extract on biofilm formation was performed on a 96-well polystyrene microtiter plate. To determine the effect of the extract on the formation of biofilms of P. aeruginosa and S. aureus, we followed the method of Mozirandi et al. 2019 [20]. The extract was serially diluted to give a final concentration range of 100-0.4 µg/mL in the wells. A 100 µL volume of numerically standardized inoculum was dispensed into each of 6 wells of the 96 well plate (Greiner 96-well plates, Sigma-Aldrich, Germany) containing an equal volume of variable concentrations of the extract. Some wells of the plate contained 200 µL of uninoculated medium only, to serve as sterility controls. Wells that had uninoculated medium and extract were also included to correct for background staining. In some wells in the plate, each bacterial strain (200 µL) was also inoculated without a plant extract and was considered a positive control for biofilm formation. The plate was incubated for 72 hours at 37 °C in a nonshaking Lab Doctor Mini Incubator (MID SCI, USA). After incubation the contents of each well were decanted into a discard container, and the plate was washed three times with sterile phosphate-buffered saline (pH=7.2) to remove free-floating non-adherent cells. The plates were then inverted and blotted on an absorbent paper towel and allowed to air dry in a sterile environment for 15 minutes [25]. The remaining attached bacteria were heat-fixed by heating the plate at 60 °C for 1 hour. The adherent biofilm layer formed in each microtiter plate well was quantified using crystal violet staining. To stain the adherent bacteria, 200 µL of 0.1% crystal violet stain was added to each well and the plate was covered and incubated at room temperature for 20 minutes. After incubation, excess stain was rinsed off by decantation. To remove the unbound dye, the plate was washed three times with distilled water and left to air dry overnight at room temperature. Each of the wells of the plate was filled with 200 µL of 95% ethanol to solubilize the biofilm-associated crystal violet dye from the cells. Optical density (OD) of the stained adherent bacteria was determined with an absorbance microplate reader using EL×800 Tecan Genios-Pro microplate reader (Grödig, Austria) at wavelength of 590 nm.
The percentage inhibition of biofilm formation was calculated using the formula in Equation 1:
1. % Biofilm inhibition=[(AB-EF)÷AB]×100 [26]
where AB is the optical density of the growth control and EF is the optical density of the sample.
Determination of the effect of the extract on mature biofilm
The ability of the extract to disrupt the mature biofilm of P. aeruginosa and S. aureus was evaluated using the methods of 2019 [20]. P. aeruginosa and S. aureus cells were cultured overnight and numerically standardized as above to inoculum densities of 5×108 CFU/mL. A volume of 100 µL of standardized cells was added to each of 6 wells of a 96-well polystyrene plate (Greiner 96-well plates, Sigma-Aldrich) and the plate was incubated for 72 hours at 37 °C in a non-shaking Lab Doctor Mini Incubator (MID SCI, USA). Cells were incubated in the absence of extracts to allow the formation of biofilms. After incubation, the contents of the wells were aspirated and non-adherent cells gently removed by washing the plate 3 times with buffered phosphate saline. Serial dilution of the extracts was prepared in a range of 0.4-100 µg/mL and added to the wells with the preformed biofilm. After the different concentrations of extracts had been added to the preformed biofilm, the plate was then incubated for 24 hours at 37 °C without shaking in the Lab Doctor Mini Incubator (MID SCI, USA). Some wells of the plate contained 200 µL of uninoculated medium only, to serve as sterility controls. Wells with uninoculated medium and extract were also included to correct for background staining.
Disruption of the biofilm structure was determined by crystal violet staining, as described above. To stain the adherent bacteria, 200 µL of 0.1% crystal violet stain was added to each well, and the plate was covered and incubated at room temperature for 20 minutes. After incubation, excess stain was rinsed off by decantation. To remove the unbound dye, the plate was washed three times with distilled water and left to air dry overnight at room temperature. Each well of the plate was filled with 200 µL of 95% ethanol to solubilise the biofilm associated crystal violet dye from the cells. OD of the stained adherent bacteria was determined using an EL×800 Tecan Genios-Pro microplate reader (Grödig, Austria) at a wavelength of 590 nm and was proportional to the biofilm that had not been disrupted by the extract. These values were then used to determine the amount of biofilm that the different concentrations of the extract used had actually destroyed.
Inhibition of metabolic activity on bacteria in biofilms
The effect of acetone extract from V. adoensis on the metabolic activity of preformed biofilms of P. aeruginosa and S. aureus was measured as described by [27] with modifications. Numerically standardized P. aeruginosa and S. aureus cells with a volume of 100 µL were added to each well of a 96-well microplate and incubated for 72 hours at 37 °C in a non-shaking incubator in the absence of extract to allow for biofilm formation as described above. Preformed biofilms were washed twice with sterile phosphate-buffered saline (PBS) and 100 µL of each extract concentration ranging from 100 to 0.4 µg/mL was added into the wells. The plates were incubated without shaking for 24 hours at 37 °C. After incubation, 50 µL of iodonitrotetrazolium (TTC, Merck, Germany) solution was added. The plate was further incubated at 37 °C for 5 hours. The absorbance of the plate was read with a microplate reader at 492 nm (Stat Fax 2100, Awareness Technology, Inc., Ramsey, Minnesota, USA). The control wells were the same as those described in the inhibition of biofilm formation assay. The inhibition of biofilm metabolic activity in the presence of different extract concentrations was calculated employing the formula described in Equation 1.
Effects of extracts on detachment of biofilms
The effect of the extracts on the detachment of P. aeruginosa and S. aureus biofilm cells was evaluated using the method of Davies et al. with adaptations [28]. P. aeruginosa and S. aureus cells were cultured overnight, and the cells were standardized as previously described to create inoculum densities of 5×108 CFU/mL. The standardized cells were incubated at 37 °C for 72 hours without agitation in the presence and absence of extract to allow for biofilm formation. The process of allowing biofilm formation by the cells was done as described in the inhibition of biofilm formation assay. However, to evaluate the effect of the extract on biofilm detachment, the step of decanting the well contents was preceded by the addition of 3 µL of sodium dodecyl sulphate (safety data sheets [SDS]; 10%) to each well, and the mixture was incubated for 30 minutes. After incubation the suspended culture was then discarded and the plate was washed twice with PBS. To stain the adherent bacteria, 200 µL of 0.1% crystal violet stain was added to each well and the plate was covered and incubated at room temperature for 20 minutes. After incubation, excess stain was rinsed off by decantation. To remove the unbound dye, the plate was washed three times with distilled water and left to air dry overnight at room temperature. Each well of the plate was filled with 200 µL of 95% ethanol to solubilise the biofilm associated crystal violet dye from the cells. OD of the stained adherent bacteria was determined using an EL×800 Tecan Genios-Pro microplate reader (Grödig, Austria) at wavelength of 590 nm and was proportional to the biofilm that remained attached to the plate following exposure to SDS and different concentrations of V. adoensis. These values were used to determine the amount of biofilm that had detached from the plate by combining V. adoensis leaf extract and SDS (10%).
UPLC-MS analysis of the acetone extract of V. adoensis
Analysis of some nonvolatile phytochemicals in the acetone extract of V. adoensis was performed by UPLC-MS and was performed according to the method of a previous study [29]. A Waters Acquity UPLC system from Waters Corporation (Milford, MA, USA) with an Acquity BEH C18 column (2.1 mm × 100 mm, particle size 1.7 μm) incorporating a binary pump, vacuum degasser, autosampler, column oven, and Micromass Xevo tandem quadrupole mass spectrometric detector (QTOF Xevo G2; Waters micromass, Manchester, UK) equipped with ESI (negative) probe. Gradient elution was used in the UPLC-MS analysis. A constant flow rate of 0.1 mL/min was used throughout the analysis at injection volumes of 10 μL. Gradient parameters were adjusted by systematically changing the percentage organic modifier at initial conditions, or the isocratic hold period at initial conditions, and/or gradient steepness. Mobile phase A was 7.5% formic acid in water, and mobile phase B was acetonitrile. The gradient started with 1% B, isocratically for 1 min followed by a linear increase to 28% at 22 min, 40% at 22.5 min, and 100% at 23 min. Column clean-up was done for 1 min at 100% B then followed by re-equilibration for 4 min at a total run time of 29 min. Electrospray mass spectra data were recorded on a negative ionisation mode for a mass range of 100 m/z to 1500 m/z at a collision energy of 50 V. Low injection volumes were used to provide responses for the main constituents within the linear range. The instrument was operated in negative ionization mode. Calibration of the instrument was done using a sodium formate solution. Data were acquired in resolution mode and mass spectrometry (MS)/MS scanning mode and processed using MassLynx software, version 4.1 (Waters). For MS/MS experiments, the trap collision energy was set to obtain sufficient fragmentation for selected precursor ions (30/45 V). The eluent was split 3:1 prior to introduction into the ionization chamber. The injection volume was 10 μL, and UV–vis spectra were acquired over 220–400 nm at 20 Hz. The accurate mass and composition for the precursor and fragment ions were determined using MassLynx software, version 4.1 (Waters) incorporated with the instrument. Using a non-targeted approach, gradient parameters were utilized by systematically adjusting solvents to determine as many compounds as possible. Using fragmentation data and available databases such as KNApSAcK metabolite database [30] and MOTO database [31] possible compounds that may be found in the V. adoensis extracts were proposed.
Statistical analyses
Graphical and statistical analyses were performed using GraphPad Prism software, version 5. Data were expressed as Mean±SD. Statistically significant differences between various means of controls and tests were analyzed using one-way analysis of variance (ANOVA) using Dunnett’s multiple comparison post-test with a P of 0.05.
Results
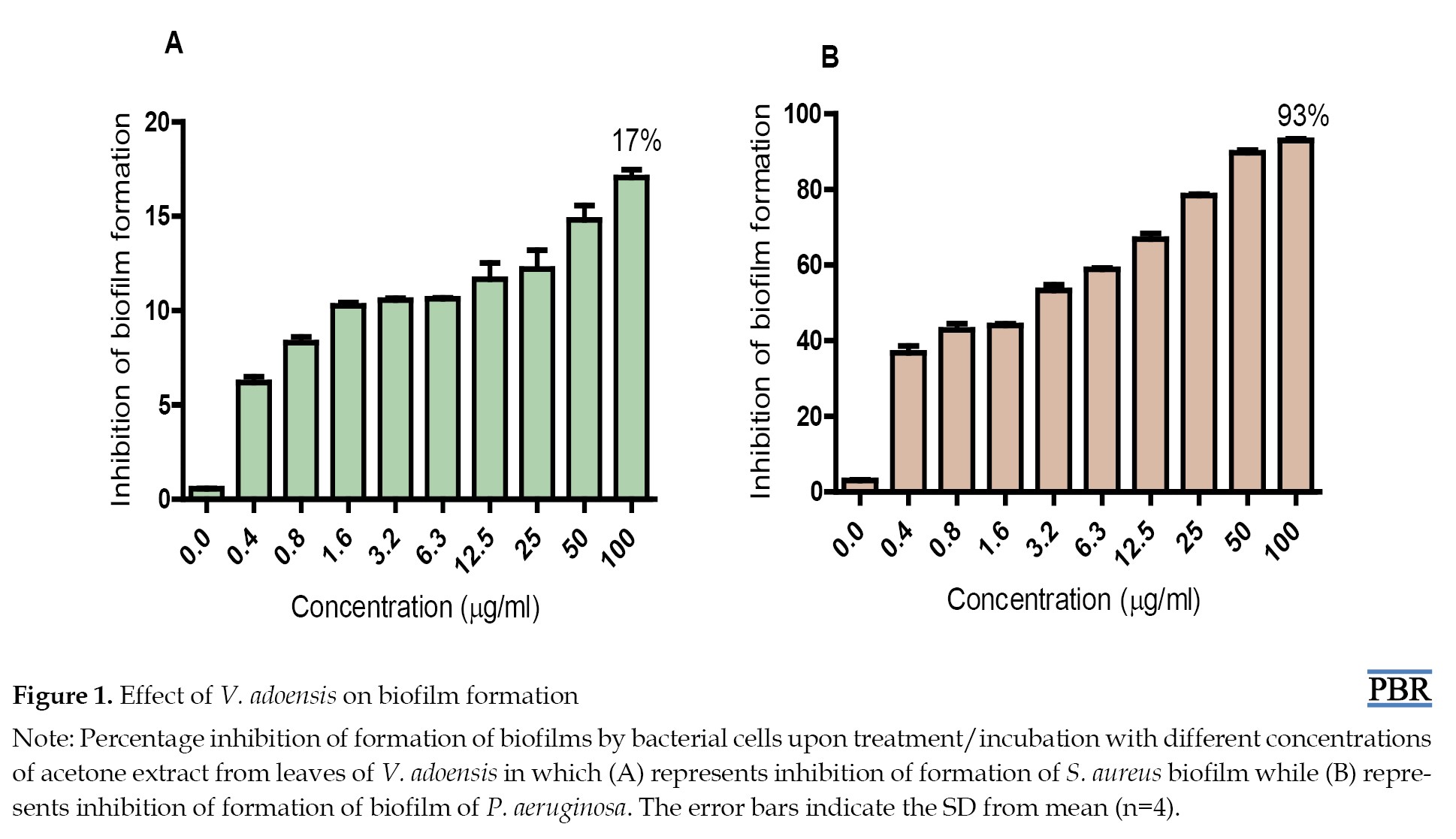
Effect of extract on biofilm formation
The effect on biofilm formation by P. aeruginosa and S. aureus was studied after 72 hours of exposure of the test strains to different concentrations of V. adoensis acetone extract. The results show that all the tested concentrations of the extract had a concentration-dependent inhibitory effect on biofilm formation by S. aureus (Figure 1A) and P.
An increase in microbial infections has increased morbidity and mortality in healthcare settings [1]. The rise of antibiotic resistance in bacteria is reported to cause an increase in nosocomial infections each year [2]. Biofilms are estimated to be responsible for more than 65% of nosocomial infections and 80% of all microbial infections [3, 4]. A biofilm is an assemblage of surface-associated microbial cells that are enclosed in an extracellular polymeric substance matrix [5]. Biofilm-related infections are difficult to eradicate as there is an increase in antimicrobial tolerance in biofilms compared to planktonic cells [6, 7]. The ability of cells in a biofilm to exert higher resistance to antimicrobials can be attributed to inactivation and reduction of penetration of active antibiotic compounds into the inner layers of the biofilms and persister cells. The increased resistance of cells in biofilm form can also be attributed to the slow growth rate of cells in biofilms and their altered metabolism, oxygen gradients, and extracellular biofilm matrix [4]. Pseudomonas aeruginosa and Staphylococcus aureus are among the most common pathogens found in biofilms in a healthcare setting [7]. These two bacterial species have been significantly identified in nosocomial infections, which have become more rampant and deadly [8]. P. aeruginosa and S. aureus are characterized by potential drug resistance mechanisms, and of major concern is their ability to form biofilms [9]. Antibiotic treatment of biofilm-related infections often provided relief from the symptoms of the infection but may not necessarily cure the infection, mainly due to the biofilms that act as reservoirs for disease recurrence [10]. Biofilms can have important negative effects on human health, for example, P. aeruginosa forms biofilms in the lungs of patients with cystic fibrosis, leading to chronic inflammation and lung tissue damage [7]. The biofilms of P. aeruginosa also cause the persistence of chronic wound infections [11]. Biofilms of S. aureus are responsible for S. aureus-related implant infections [12].
Although biofilms often resist to conventional antibiotic therapy, there are promising alternatives considering the reported efficacy of various plant metabolites against bacteria in biofilms. Phytochemicals from medicinal plants that show antimicrobial activities have the potential to be alternative antimicrobial agents, as their structures differ from those of the more studied microbial sources [13, 14]. Therefore, their mode of action may very likely differ [15]. Vernonia adoensis is a commonly used African ethnomedicinal plant that has been proven to have antimicrobial activity [16, 17]. Screening active compounds from some Vernonia species has led to the discovery of new lead compounds which have efficient protection and treatment roles, including anticancer [18], antiplasmodial [19] and antibacterial [20] activity. Primary phytochemical screening of V. adoensis collected from Kenya has shown some important pharmacological phytochemicals, such as phenols, saponnins, flavonoids, glycosides, tannins, alkaloids, and terpenoids [21]. The composition of phytochemical components varies with geographical location and harvesting conditions [22, 23]. Therefore, the current study aimed to determine the possible chemical components of phytoconstituents from V. adoensis leaves collected from Zimbabwe using gas chromatography–mass spectrometry (GC-MS) and ultra-performance liquid chromatography-mass spectroscopy (UPLC-MS) techniques, and to evaluate the antibiofilm activity of the phytochemicals.
Materials and Methods
Plant material collection
V. adoensis was collected in Centenary, (Geographic coordinates, Latitude: 16º43’22’’ S, Longitude: 31º06’52’’ E, elevation above sea level: 1156 m) Mashonaland Central Province of Zimbabwe. Plant collection was performed in March 2018. Authentication and classification of the plant were performed by a taxonomist from the National Herbarium and Botanic Garden (Harare, Zimbabwe). Herbarium samples C1 E7, were kept at the National Herbarium and Botanic Garden (Harare, Zimbabwe) and the Department of Biochemistry, University of Zimbabwe.
Preparation of acetone extract
The leaf acetone extract of V. adoensis has been reported to have an inhibitory effect on the growth of both P. aeruginosa and S. aureus [17]; therefore, the acetone extract was used to evaluate the effect of V. adoensis on biofilms of S. aureus and P. aeruginosa. The extract was prepared using Mozirandi et al. 2019 [20].
V. adoensis leaves were washed under running tap water and dried in the oven at 40 oC. The dried leaves were pounded in a clean mortar and further ground in a two-speed blender (Cole Parmer Instrument CO., Vernon Hills, USA) to obtain a fine powder to which 10 mL of acetone (Sigma-Aldrich, Taufkirchen, Germany) was added to each 1 g of plant material to extract phytochemicals by maceration [24]. The extract was then filtered using Whatman’s no one filter paper (Sigma-Aldrich, Taufkirchen, Germany). The filtrate was concentrated to dryness by evaporation at room temperature in a fume hood with air stream. The dry extract was stored at 4 ºC for further use.
Bacteria and culture conditions
S. aureus (ATCC 9144) and P. aeruginosa (ATCC 27853), obtained from the Division of Microbiology, Department of Biological Sciences, University of Botswana were separately inoculated in TSB supplemented with 1% glucose and cultured overnight at 37 ºC in a shaking incubator at 120 rpm (SI300 Incubated shaker, Jeiotech, Korea). Cells were centrifuged at 3500 rpm for 4 min on a Hettich Rotofix 32 centrifuge (Tuttlingen, Germany). The pellet was then suspended in fresh media and a 0.5 McFarland Standard was used to create inoculum densities of 5×108 CFU/mL.
Effect of the extract on biofilm formation
Evaluation of the effect of the extract on biofilm formation was performed on a 96-well polystyrene microtiter plate. To determine the effect of the extract on the formation of biofilms of P. aeruginosa and S. aureus, we followed the method of Mozirandi et al. 2019 [20]. The extract was serially diluted to give a final concentration range of 100-0.4 µg/mL in the wells. A 100 µL volume of numerically standardized inoculum was dispensed into each of 6 wells of the 96 well plate (Greiner 96-well plates, Sigma-Aldrich, Germany) containing an equal volume of variable concentrations of the extract. Some wells of the plate contained 200 µL of uninoculated medium only, to serve as sterility controls. Wells that had uninoculated medium and extract were also included to correct for background staining. In some wells in the plate, each bacterial strain (200 µL) was also inoculated without a plant extract and was considered a positive control for biofilm formation. The plate was incubated for 72 hours at 37 °C in a nonshaking Lab Doctor Mini Incubator (MID SCI, USA). After incubation the contents of each well were decanted into a discard container, and the plate was washed three times with sterile phosphate-buffered saline (pH=7.2) to remove free-floating non-adherent cells. The plates were then inverted and blotted on an absorbent paper towel and allowed to air dry in a sterile environment for 15 minutes [25]. The remaining attached bacteria were heat-fixed by heating the plate at 60 °C for 1 hour. The adherent biofilm layer formed in each microtiter plate well was quantified using crystal violet staining. To stain the adherent bacteria, 200 µL of 0.1% crystal violet stain was added to each well and the plate was covered and incubated at room temperature for 20 minutes. After incubation, excess stain was rinsed off by decantation. To remove the unbound dye, the plate was washed three times with distilled water and left to air dry overnight at room temperature. Each of the wells of the plate was filled with 200 µL of 95% ethanol to solubilize the biofilm-associated crystal violet dye from the cells. Optical density (OD) of the stained adherent bacteria was determined with an absorbance microplate reader using EL×800 Tecan Genios-Pro microplate reader (Grödig, Austria) at wavelength of 590 nm.
The percentage inhibition of biofilm formation was calculated using the formula in Equation 1:
1. % Biofilm inhibition=[(AB-EF)÷AB]×100 [26]
where AB is the optical density of the growth control and EF is the optical density of the sample.
Determination of the effect of the extract on mature biofilm
The ability of the extract to disrupt the mature biofilm of P. aeruginosa and S. aureus was evaluated using the methods of 2019 [20]. P. aeruginosa and S. aureus cells were cultured overnight and numerically standardized as above to inoculum densities of 5×108 CFU/mL. A volume of 100 µL of standardized cells was added to each of 6 wells of a 96-well polystyrene plate (Greiner 96-well plates, Sigma-Aldrich) and the plate was incubated for 72 hours at 37 °C in a non-shaking Lab Doctor Mini Incubator (MID SCI, USA). Cells were incubated in the absence of extracts to allow the formation of biofilms. After incubation, the contents of the wells were aspirated and non-adherent cells gently removed by washing the plate 3 times with buffered phosphate saline. Serial dilution of the extracts was prepared in a range of 0.4-100 µg/mL and added to the wells with the preformed biofilm. After the different concentrations of extracts had been added to the preformed biofilm, the plate was then incubated for 24 hours at 37 °C without shaking in the Lab Doctor Mini Incubator (MID SCI, USA). Some wells of the plate contained 200 µL of uninoculated medium only, to serve as sterility controls. Wells with uninoculated medium and extract were also included to correct for background staining.
Disruption of the biofilm structure was determined by crystal violet staining, as described above. To stain the adherent bacteria, 200 µL of 0.1% crystal violet stain was added to each well, and the plate was covered and incubated at room temperature for 20 minutes. After incubation, excess stain was rinsed off by decantation. To remove the unbound dye, the plate was washed three times with distilled water and left to air dry overnight at room temperature. Each well of the plate was filled with 200 µL of 95% ethanol to solubilise the biofilm associated crystal violet dye from the cells. OD of the stained adherent bacteria was determined using an EL×800 Tecan Genios-Pro microplate reader (Grödig, Austria) at a wavelength of 590 nm and was proportional to the biofilm that had not been disrupted by the extract. These values were then used to determine the amount of biofilm that the different concentrations of the extract used had actually destroyed.
Inhibition of metabolic activity on bacteria in biofilms
The effect of acetone extract from V. adoensis on the metabolic activity of preformed biofilms of P. aeruginosa and S. aureus was measured as described by [27] with modifications. Numerically standardized P. aeruginosa and S. aureus cells with a volume of 100 µL were added to each well of a 96-well microplate and incubated for 72 hours at 37 °C in a non-shaking incubator in the absence of extract to allow for biofilm formation as described above. Preformed biofilms were washed twice with sterile phosphate-buffered saline (PBS) and 100 µL of each extract concentration ranging from 100 to 0.4 µg/mL was added into the wells. The plates were incubated without shaking for 24 hours at 37 °C. After incubation, 50 µL of iodonitrotetrazolium (TTC, Merck, Germany) solution was added. The plate was further incubated at 37 °C for 5 hours. The absorbance of the plate was read with a microplate reader at 492 nm (Stat Fax 2100, Awareness Technology, Inc., Ramsey, Minnesota, USA). The control wells were the same as those described in the inhibition of biofilm formation assay. The inhibition of biofilm metabolic activity in the presence of different extract concentrations was calculated employing the formula described in Equation 1.
Effects of extracts on detachment of biofilms
The effect of the extracts on the detachment of P. aeruginosa and S. aureus biofilm cells was evaluated using the method of Davies et al. with adaptations [28]. P. aeruginosa and S. aureus cells were cultured overnight, and the cells were standardized as previously described to create inoculum densities of 5×108 CFU/mL. The standardized cells were incubated at 37 °C for 72 hours without agitation in the presence and absence of extract to allow for biofilm formation. The process of allowing biofilm formation by the cells was done as described in the inhibition of biofilm formation assay. However, to evaluate the effect of the extract on biofilm detachment, the step of decanting the well contents was preceded by the addition of 3 µL of sodium dodecyl sulphate (safety data sheets [SDS]; 10%) to each well, and the mixture was incubated for 30 minutes. After incubation the suspended culture was then discarded and the plate was washed twice with PBS. To stain the adherent bacteria, 200 µL of 0.1% crystal violet stain was added to each well and the plate was covered and incubated at room temperature for 20 minutes. After incubation, excess stain was rinsed off by decantation. To remove the unbound dye, the plate was washed three times with distilled water and left to air dry overnight at room temperature. Each well of the plate was filled with 200 µL of 95% ethanol to solubilise the biofilm associated crystal violet dye from the cells. OD of the stained adherent bacteria was determined using an EL×800 Tecan Genios-Pro microplate reader (Grödig, Austria) at wavelength of 590 nm and was proportional to the biofilm that remained attached to the plate following exposure to SDS and different concentrations of V. adoensis. These values were used to determine the amount of biofilm that had detached from the plate by combining V. adoensis leaf extract and SDS (10%).
UPLC-MS analysis of the acetone extract of V. adoensis
Analysis of some nonvolatile phytochemicals in the acetone extract of V. adoensis was performed by UPLC-MS and was performed according to the method of a previous study [29]. A Waters Acquity UPLC system from Waters Corporation (Milford, MA, USA) with an Acquity BEH C18 column (2.1 mm × 100 mm, particle size 1.7 μm) incorporating a binary pump, vacuum degasser, autosampler, column oven, and Micromass Xevo tandem quadrupole mass spectrometric detector (QTOF Xevo G2; Waters micromass, Manchester, UK) equipped with ESI (negative) probe. Gradient elution was used in the UPLC-MS analysis. A constant flow rate of 0.1 mL/min was used throughout the analysis at injection volumes of 10 μL. Gradient parameters were adjusted by systematically changing the percentage organic modifier at initial conditions, or the isocratic hold period at initial conditions, and/or gradient steepness. Mobile phase A was 7.5% formic acid in water, and mobile phase B was acetonitrile. The gradient started with 1% B, isocratically for 1 min followed by a linear increase to 28% at 22 min, 40% at 22.5 min, and 100% at 23 min. Column clean-up was done for 1 min at 100% B then followed by re-equilibration for 4 min at a total run time of 29 min. Electrospray mass spectra data were recorded on a negative ionisation mode for a mass range of 100 m/z to 1500 m/z at a collision energy of 50 V. Low injection volumes were used to provide responses for the main constituents within the linear range. The instrument was operated in negative ionization mode. Calibration of the instrument was done using a sodium formate solution. Data were acquired in resolution mode and mass spectrometry (MS)/MS scanning mode and processed using MassLynx software, version 4.1 (Waters). For MS/MS experiments, the trap collision energy was set to obtain sufficient fragmentation for selected precursor ions (30/45 V). The eluent was split 3:1 prior to introduction into the ionization chamber. The injection volume was 10 μL, and UV–vis spectra were acquired over 220–400 nm at 20 Hz. The accurate mass and composition for the precursor and fragment ions were determined using MassLynx software, version 4.1 (Waters) incorporated with the instrument. Using a non-targeted approach, gradient parameters were utilized by systematically adjusting solvents to determine as many compounds as possible. Using fragmentation data and available databases such as KNApSAcK metabolite database [30] and MOTO database [31] possible compounds that may be found in the V. adoensis extracts were proposed.
Statistical analyses
Graphical and statistical analyses were performed using GraphPad Prism software, version 5. Data were expressed as Mean±SD. Statistically significant differences between various means of controls and tests were analyzed using one-way analysis of variance (ANOVA) using Dunnett’s multiple comparison post-test with a P of 0.05.
Results
Effect of extract on biofilm formation
The effect on biofilm formation by P. aeruginosa and S. aureus was studied after 72 hours of exposure of the test strains to different concentrations of V. adoensis acetone extract. The results show that all the tested concentrations of the extract had a concentration-dependent inhibitory effect on biofilm formation by S. aureus (Figure 1A) and P.
aeruginosa (Figure 1B). However, no concentration of the extract was observed that completely inhibited formation of biofilm by both test strains. The most susceptible bacteria were P. aeruginosa, in which the highest concentration (100 µg/mL) of extract inhibited the formation of biofilm of the bacteria by 93% while S. aureus biofilm formation was inhibited by 17% (Figures 1A and 1B).
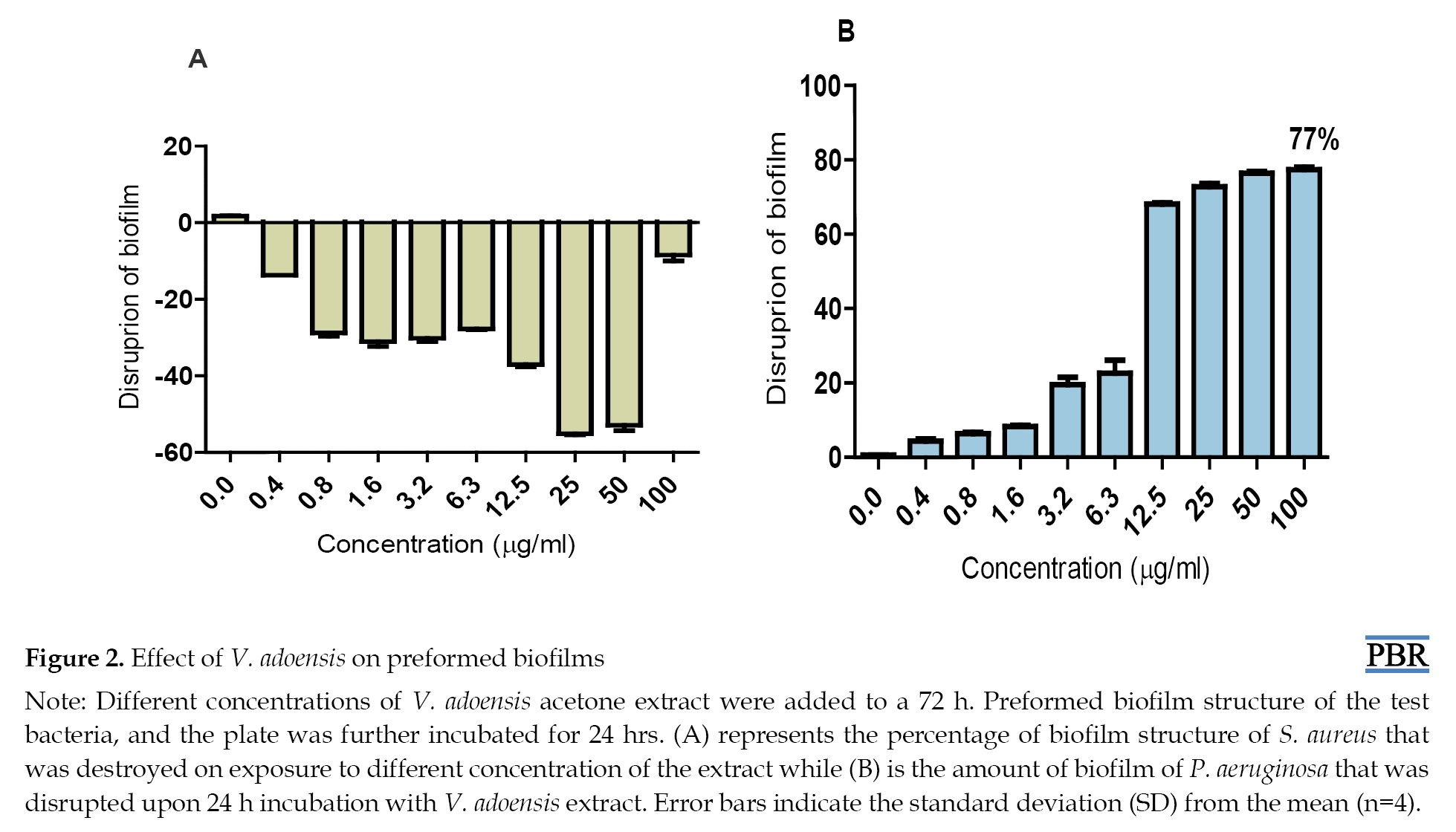
Effect of extract on mature biofilm
P. aeruginosa and S. aureus were allowed to form biofilms for 72 hours in the absence of V. adoensis acetone extract after which the extract was then incorporated and its effect evaluated after 24-hr. incubation. The results showed that as the concentration of the extract increased, an increased disruption of preformed biofilm of P. aeruginosa was observed. Although no concentration of the extract was observed which managed to destroy the biofilm structure of P. aeruginosa, a total of 77% of biofilm of the bacteria was disrupted after exposure to the highest (100 µg/mL) concentration of the extract (Figure 2B).
Effect of extract on mature biofilm
P. aeruginosa and S. aureus were allowed to form biofilms for 72 hours in the absence of V. adoensis acetone extract after which the extract was then incorporated and its effect evaluated after 24-hr. incubation. The results showed that as the concentration of the extract increased, an increased disruption of preformed biofilm of P. aeruginosa was observed. Although no concentration of the extract was observed which managed to destroy the biofilm structure of P. aeruginosa, a total of 77% of biofilm of the bacteria was disrupted after exposure to the highest (100 µg/mL) concentration of the extract (Figure 2B).
The results of this study also showed that none of the concentrations of V. adoensis used in this study disrupted preformed biofilms of S. aureus (Figure 2A).
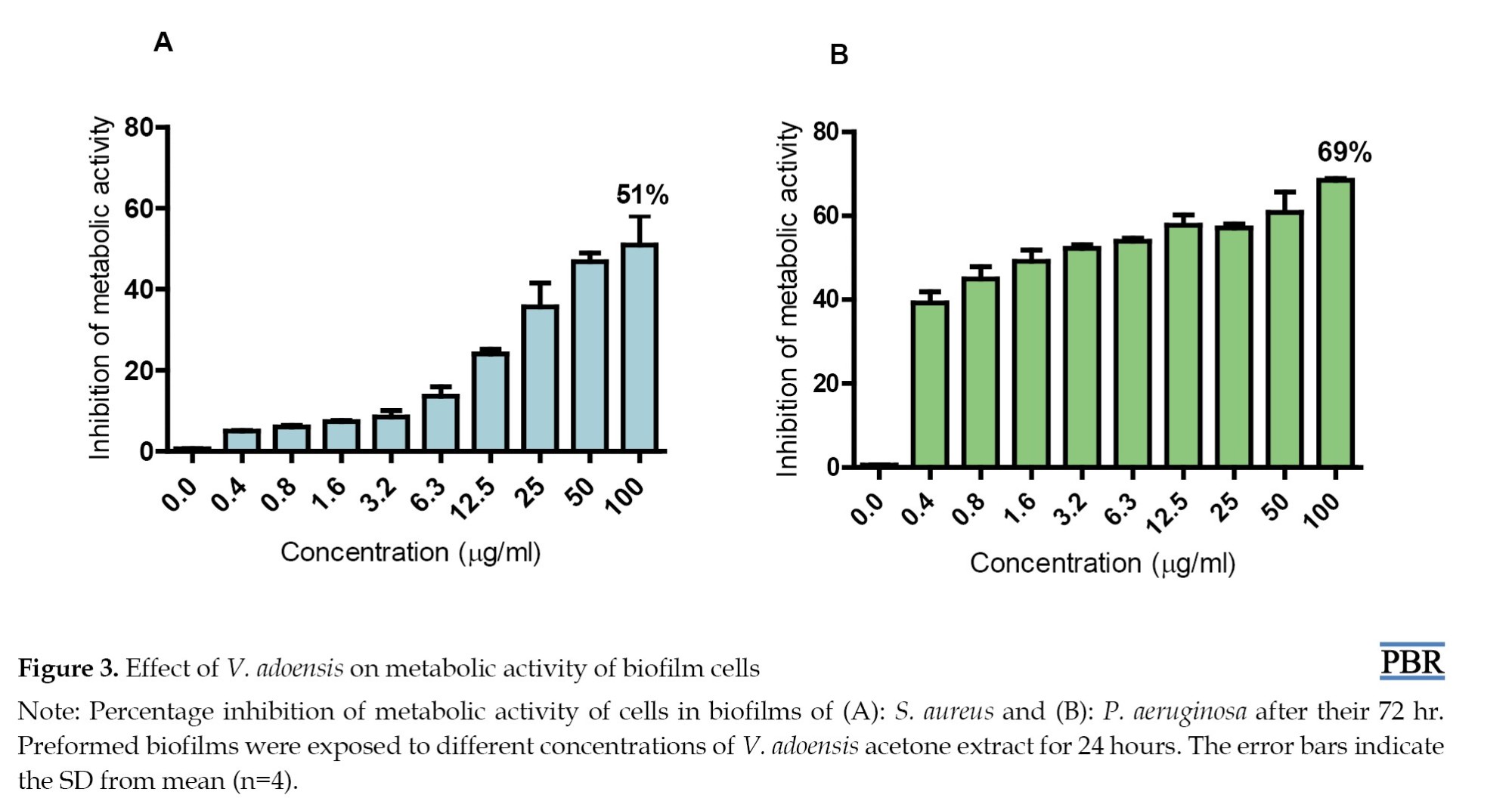
Inhibition of metabolic activity on bacteria in biofilms
V. adoensis acetone extract was added to a 72-hr mature biofilm of S. aureus and P. aeruginosa and incubated for 24 hours to evaluate its effect on the metabolic activity of the respective bacteria in the biofilm structure. The results showed that the extract has an inhibitory effect on the metabolic activity of the bacteria in biofilms of both S. aureus and P. aeruginosa (Figures 3A and 3B).
Inhibition of metabolic activity on bacteria in biofilms
V. adoensis acetone extract was added to a 72-hr mature biofilm of S. aureus and P. aeruginosa and incubated for 24 hours to evaluate its effect on the metabolic activity of the respective bacteria in the biofilm structure. The results showed that the extract has an inhibitory effect on the metabolic activity of the bacteria in biofilms of both S. aureus and P. aeruginosa (Figures 3A and 3B).
V. adoensis extract inhibited the metabolic activity of cells in biofilm of P. aeruginosa by 69% (Figure 3B) and the metabolic activity of cells in S. aureus biofilm was inhibited by 51% (Figure 3A).
Effect of extract on the detachment of biofilm cells
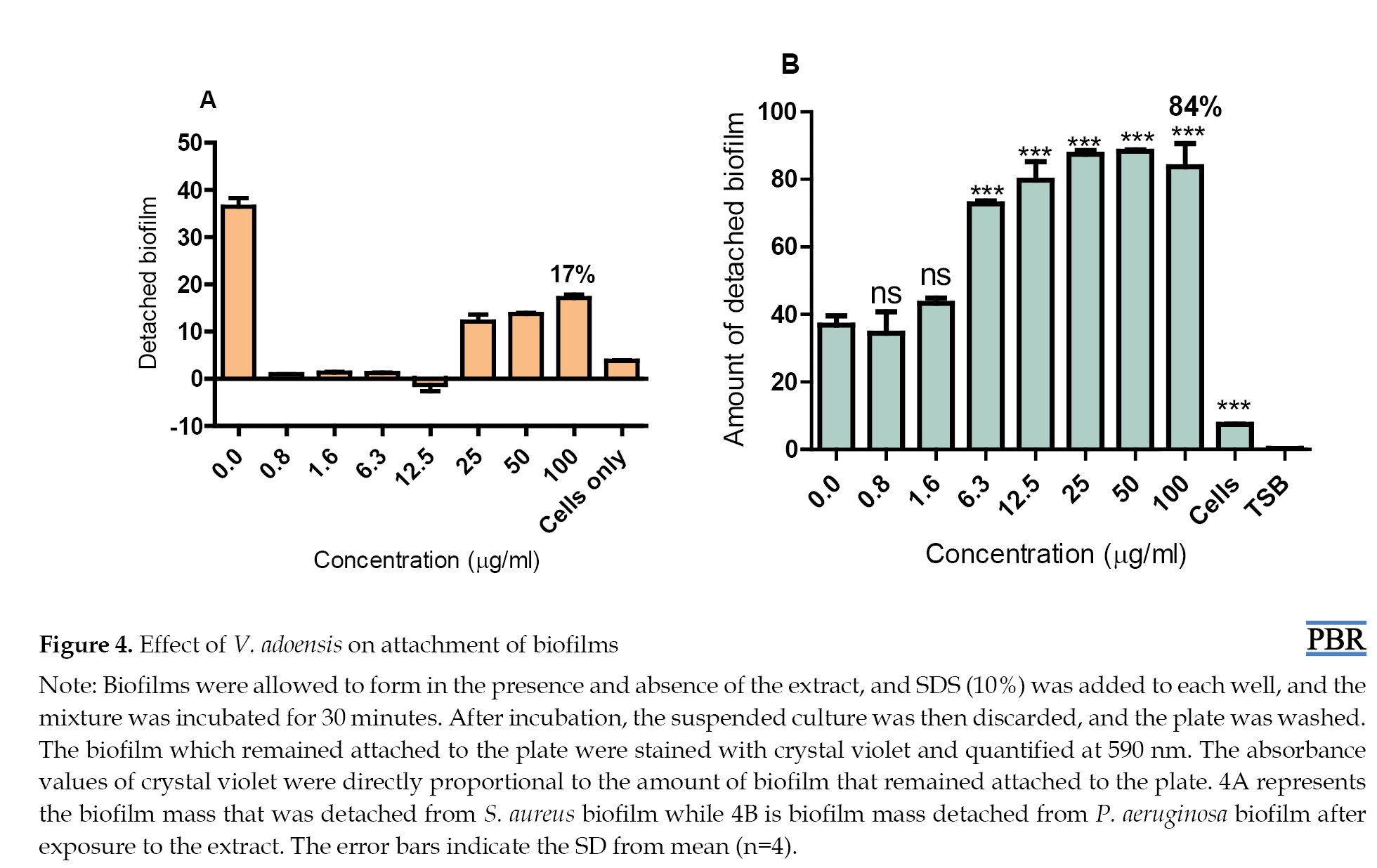
To evaluate the effect of V. adoensis on attachment of biofilm cells, we quantified the amount of biofilm after treatment with SDS in biofilms formed in the presence and absence of different concentrations of V. adoensis acetone extract. Unexposed cells represented detached biofilm that had been formed from cells that had neither been exposed to the extract nor SDS, while detached biofilm at 0.0 concentration is from the biofilm detached by SDS after it had been formed in the absence of the extract. The results showed that more biofilm was detached from both treated and untreated biofilms (no extract i.e. 0 µg/mL) relative to biofilm detached in cells not exposed to the surfactant (cells only). Therefore, SDS caused detachment of biofilms formed both in the presence and absence of the extract (Figure 4).
Effect of extract on the detachment of biofilm cells
To evaluate the effect of V. adoensis on attachment of biofilm cells, we quantified the amount of biofilm after treatment with SDS in biofilms formed in the presence and absence of different concentrations of V. adoensis acetone extract. Unexposed cells represented detached biofilm that had been formed from cells that had neither been exposed to the extract nor SDS, while detached biofilm at 0.0 concentration is from the biofilm detached by SDS after it had been formed in the absence of the extract. The results showed that more biofilm was detached from both treated and untreated biofilms (no extract i.e. 0 µg/mL) relative to biofilm detached in cells not exposed to the surfactant (cells only). Therefore, SDS caused detachment of biofilms formed both in the presence and absence of the extract (Figure 4).
However, the detachment efficiency of SDS for biofilm cells of P. aeruginosa biofilm formed in the presence of V. adoensis extract was higher than that for biofilm cells formed in the absence of the extract. Less than 43% of biofilm was detached from P. aeruginosa non-treated biofilm while there was significant detachment of biofilm formed in the presence of the extract starting from 6.3 µg/mL (Figure 4B). This result suggested that P. aeruginosa biofilms formed in the presence of higher concentrations of V. adoensis (6.3-100 µg/mL) were more loosely attached to the surface than the biofilm formed in the absence of the extract (i.e. at 0 µg/mL). At lower concentrations of the extract (0.8-1.6 µg/mL), no significant difference was observed in the detached cells from the untreated and treated biofilms of P. aeruginosa. This shows that lower concentrations of V. adoensis do not affect attachment of biofilms to the plate.
The results also showed that the V. adoensis extract did not enhance the detachment of biofilms of S. aureus by the surfactant; only 17% of biofilm was detached from treated biofilm, relative to 36% detached from biofilm formed in the absence of the extract (Figure 4A).
UPLC-MS analysis
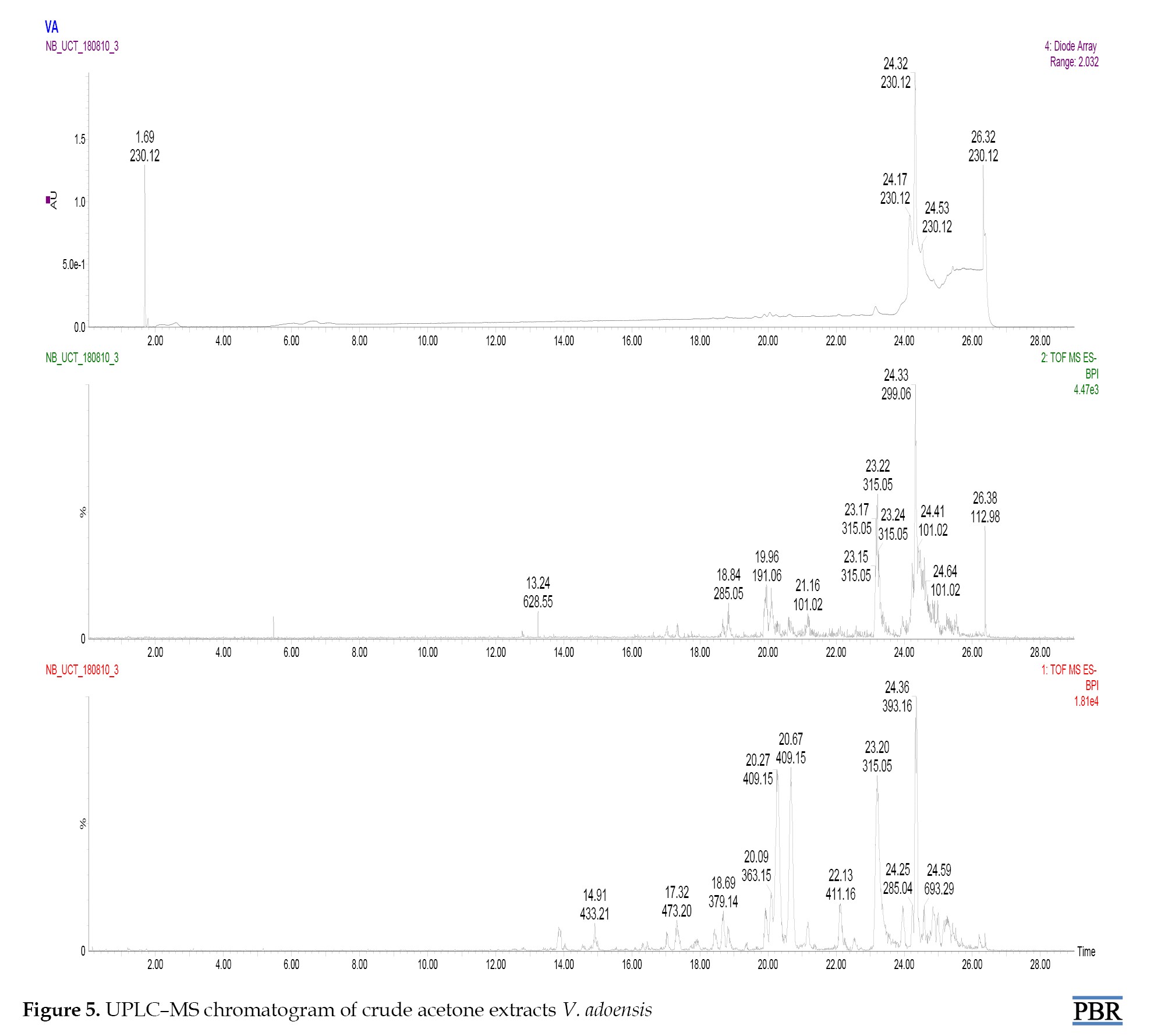
UPLC-MS analysis of V. adoensis identified several phytochemicals represented in the chromatogram (Figure 5).
The results also showed that the V. adoensis extract did not enhance the detachment of biofilms of S. aureus by the surfactant; only 17% of biofilm was detached from treated biofilm, relative to 36% detached from biofilm formed in the absence of the extract (Figure 4A).
UPLC-MS analysis
UPLC-MS analysis of V. adoensis identified several phytochemicals represented in the chromatogram (Figure 5).
The phytochemicals were tentatively characterized and identified in Table 1 presents ESI-MS/MS data and fragmentations.
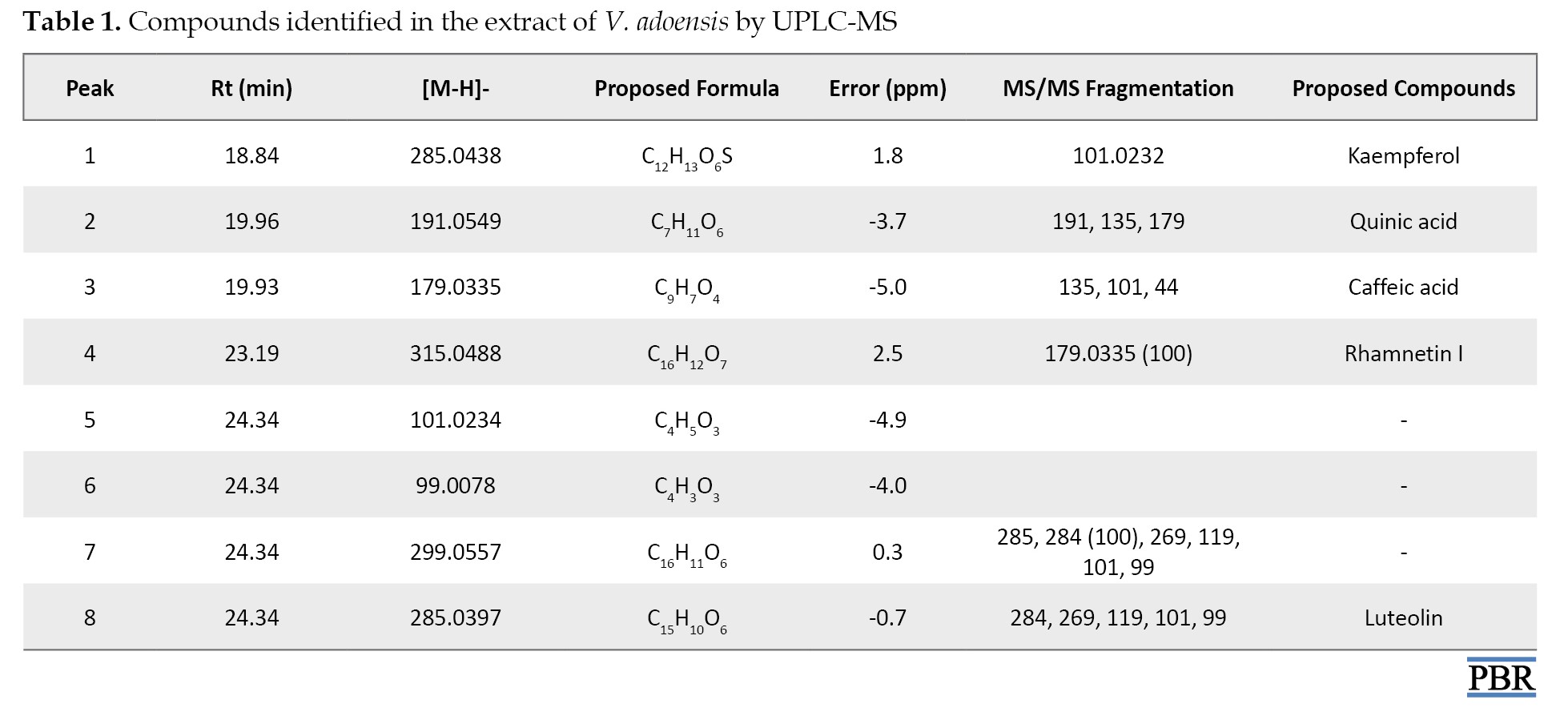
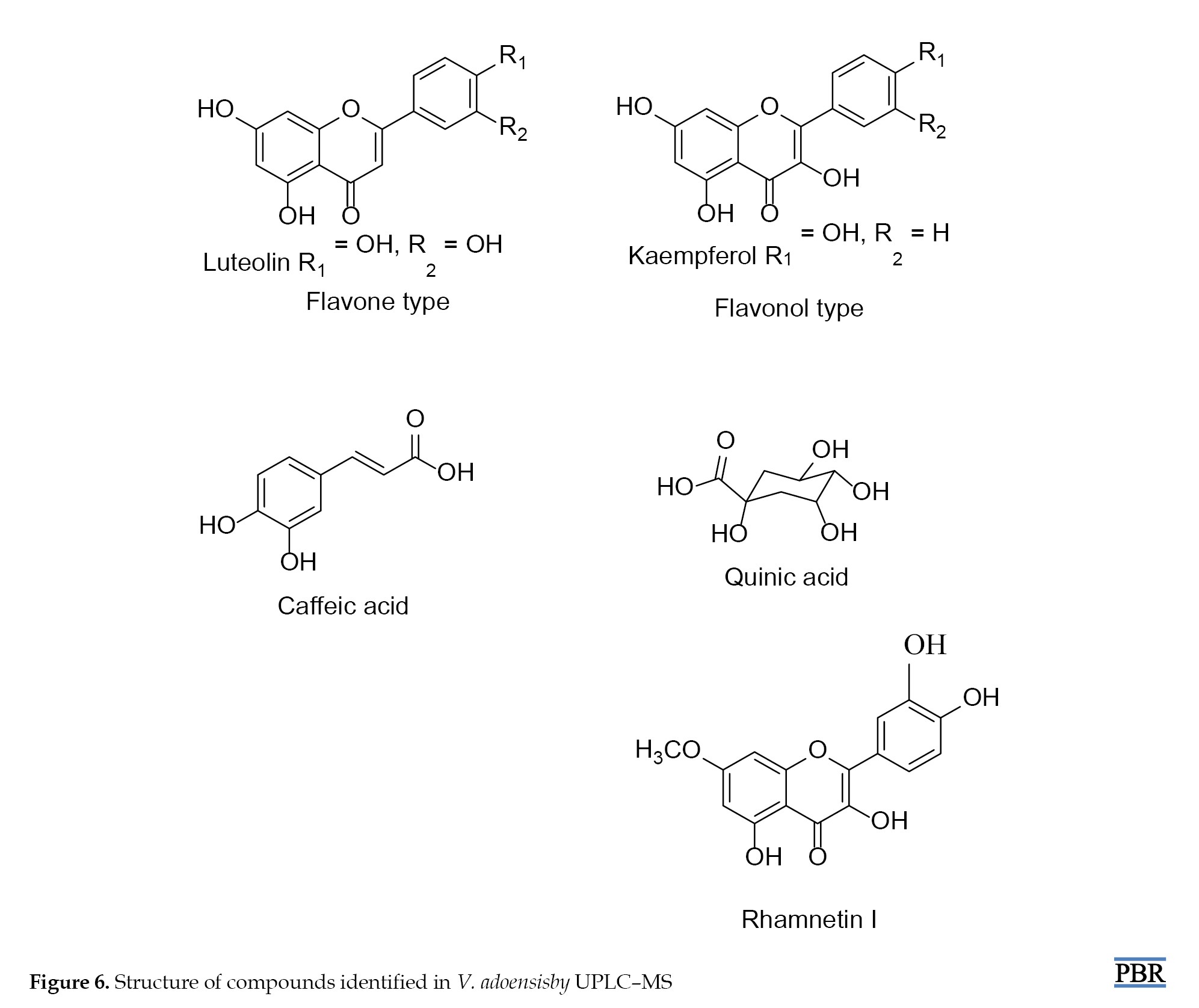
Tentatively identified compounds from V. adoensis included kaempferol, quinic acid, caffeic acid, rhamnetin I, and luteolin. Selected structures of these phytochemicals are assigned (Figure 6).
Tentatively identified compounds from V. adoensis included kaempferol, quinic acid, caffeic acid, rhamnetin I, and luteolin. Selected structures of these phytochemicals are assigned (Figure 6).
Discussion
This study aimed to investigate the effects of V. adoensis leaf extracts on S. aureus and P. aeruginosa biofilms and determine the phytochemical composition of the extract by UPLC-MS. The leaf extract displayed antibiofilm potential against formation of biofilms by both P. aeruginosa and S. aureus. Since bacterial resistance to antibiotics and their survival are associated with their ability to form biofilms [32], extracts that have the potential to decrease biofilm formation would be useful and can be used in conjunction with the antibiotics to decrease bacterial resistance. The results of this study are similar to other studies which have also been done and shown that extracts are capable of inhibiting formation of biofilms by bacteria [33, 14].
When biofilm-forming pathogens such as P. aeruginosa and S. aureus form biofilms in patients, despite the aggressive use of antibiotics, colonization is often a life-long problem leading to chronic diseases [7]. Therefore, preventing the pathogen’s transition from the planktonic state to the biofilm growth mode is the most important step for combating biofilm-associated pathogens, as the ability of pathogens to resist antibiotics is significantly enhanced 10 to 1000 times once they form biofilms [32]
Clinically, biofilm inhibitors can be used directly to reduce virulence factors from the infectious bacteria [3] or to treat biofilm along with conventional antibiotic [33]. The extract used in this study has shown the ability to destruct mature biofilms of P. aeruginosa; therefore, it may be useful in destruction or reduction of preformed biofilms of the bacteria. The weakening or destruction of the mature biomass can be followed by washout or subsequent biocidal inactivation of the detached biomass [34]. Weakening or destroying the mature biofilm will render cells susceptible to antibiotics. It was observed that all the tested concentrations of the extract significantly inhibited metabolic activity on bacteria in preformed biofilms for all tested strains. Plants contain various secondary metabolites, some of which have been found to exert a range of biological activities [35, 36]. The antibiofilm activity produced by the extracts could be due to the presence of individual phytochemicals that are active against S. aureus and P. aeruginosa biofilm or could be the result of the synergistic effect of two or more phytochemicals contained in the extract.
Analysis of plant extracts helps to determine whether the plant species contains any individual compound or group of compounds, which may substantiate its current commercial and traditional use as herbal medicine. Further phytochemical analysis also helps to determine the most appropriate methods of extracting these compounds. Chromatographic techniques contribute significantly to natural products, especially regarding identification, separation, and characterization of bioactive compounds from plant sources [37]. In this study, UPLC-MS was used and UPLC- MS can identify semipolar metabolites with advantages of high precision and short time consumed.
In this study, UPLC-MS analysis of crude acetone extract of V. adoensis revealed the presence of more than five phytochemicals, most of which had been shown to possess some pharmacological properties. For instance, kaempferol, one of the compounds identified in the study, has been shown to exhibit good antioxidant activity [38, 39], anti-inflammatory [40] and anticancer activity [41]. Kaempferol had also been previously identified from Arum palaestinum (Araceae) leaves in a similar study by Abu-reidar [42]. Another compound identified by UPLC-MS was quinic acid which has been shown to have potent anti-herpes activity [43]. Quinic acid has also been previously identified in a similar study [42]. UPLC-MS also identified the presence of caffeic acid in V. adoensis leaves and this phytocompound has been reported to be a potential anti-inflammatory and anticancer agent [44, 45]. A similar study identified caffeic acid in plant species (Hyptis suaveolens and Boerhavia diffusa leaves) [29]. Some compounds identified in the extract by UPLC-MS including rhamnetin and luteon have also been shown to possess biological activity. Rhamnetin was shown to have anti-inflammatory as well as free radical scavenging activity [46, 47]. Luteon has been shown to possess anti-inflammatory [48] and anticancer activities [49]. Similar studies had also identified rhamnetin in betula [50] while Luteolin was identified from burdock, Arctium lappa L leaves [51]. The phytochemical profiles may be used as a pharmacognostical tool for the identification of the plants and as a starting point in the construction of chromatographic fingerprints which can be useful for evaluating the quality of herbal medicines [52, 53]. The observed antibiofilm activity of the extract can be attributed to the different phytochemical constituents that have been found to present in it.
Conclusion
Leaf extracts from V. adoensis have antibiofilm activity on P. aeruginosa and S. aureus biofilms. The phytochemicals identified in V. adoensis extracts may play major roles in the reported biological activities and pharmacological properties of the plant and could lead to novel drugs with potent antibiofilm activity. Further follow-up studies are required in the future in order to elucidate the underlying mechanism of the antibiofilm activity of the extract.
Ethical Considerations
Compliance with ethical guidelines
The study was conducted according to the protocol approved by the Faculty of Science Higher Degrees Committee University of Zimbabwe, Harare, Zimbabwe (Code: Paper HD/166 of 2016).
Funding
This study was supported by the Swedish International Development Agency via the International Science Programs (ISP), and through the International Program in the Chemical Sciences Uppsala University, Uppsala, Sweden (Code: ISP IPICS: ZIM01) and the International Foundation for Sciences (IFS), Stockholm, Sweden (Code: IFS F/3413-03F). This study was supported by the Alliance for Global Health and Science (University of California, Berkeley, Unites States).
Authors' contributions
Conceptualization, study design, and project administration: Stanley Mukanganyama; Experiments: Winnie Mozirandi; Data interpretation: Chi Godloves Fru; Writing and final approval: All authors.
Conflict of interest
The authors declared no conflict of interest.
Acknowledgments
The authors acknowledge the assistance of Christopher Chapano, a taxonomist at the National Herbarium and Botanical Gardens, Harare, Zimbabwe, in authenticating the plant sample names.
References
- Ara N, Nur MH, Amran MS, Wahid MI, Ahmed M. In vitro antimicrobial and cytotoxic activities of leaves and flowers extracts from Lippia alba. Pak J Biol Sci. 2009; 12(1):87-90. [DOI:10.3923/pjbs.2009.87.90] [PMID]
- Tiwari N, Rajdev S, Mullan S. Resistance trends among Pseudomonas aeruginosa isolates in a tertiary care centre in South Gujarat. Adv Microbiol. 2017; 7(3):188-94. [Link]
- Davies D. Understanding biofilm resistance to antibacterial agents. Nat Rev Drug Discov. 2003; 2(2):114-22. [DOI:10.1038/nrd1008] [PMID]
- Romling U, Balsalobre C. Biofilm infections, their resilience to therapy and innovative treatment strategies. J Intern Med. 2012; 272(6):541-61. [DOI:10.1111/joim.12004] [PMID]
- Branda SS, González-Pastor JE, Ben-Yehuda S, Losick R, Kolter R. Fruiting body formation by Bacillus subtilis. Proc Natl Acad Sci U S A. 2001; 98(20):11621-6. [DOI:10.1073/pnas.191384198] [PMID]
- Ceri H, Olson ME, Stremick C, Read RR, Morck D, Buret A. The Calgary Biofilm Device: New technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J Clin Microbiol. 1999; 37(6):1771-6. [DOI:10.1128/JCM.37.6.1771-1776.1999] [PMID]
- Høiby N, Bjarnsholt T, Givskov M, Molin S, Ciofu O. Antibiotic resistance of bacterial biofilms. Int J Antimicrob Agents. 2010; 35(4):322-32. [DOI:10.1016/j.ijantimicag.2009.12.011] [PMID]
- Rice LB. Progress and challenges in implementing the research on ESKAPE pathogens. Infect Control Hosp Epidemiol. 2010; 31 (Suppl 1):S7-10. [DOI:10.1086/655995] [PMID]
- Donlan RM, Costerton JW. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin Microbiol Rev. 2002; 15(2):167-93. [DOI:10.1128/CMR.15.2.167-193.2002] [PMID]
- Costerton JW, Stewart PS, Greenberg EP. Bacterial biofilms: A common cause of persistent infections. Science. 1999; 284(5418):1318-22. [DOI:10.1126/science.284.5418.1318] [PMID]
- James GA, Swogger E, Wolcott R, Pulcini Ed, Secor P, Sestrich J, et al. Biofilms in chronic wounds. Wound Repair Regen. 2008; 16(1):37-44. [DOI:10.1111/j.1524-475X.2007.00321.x] [PMID]
- Arduino JM, Kaye KS, Reed SD, Peter SA, Sexton DJ, Chen LF, et al. Staphylococcus aureus infections following knee and hip prosthesis insertion procedures. Antimicrob Resist Infect Control. 2015; 4:13. [DOI:10.1186/s13756-015-0057-4] [PMID]
- Araby E, El-Tablawy SY. Inhibitory effects of rosemary (Rosemarinus officinalis L.) essential oil on pathogenicity of irradiated and non-irradiated Pseudomonas aeruginosa. J Photochem Photobiol B. 2016; 159:24-32. [DOI:10.1016/j.jphotobiol.2016.02.024] [PMID]
- Kim YG, Lee JH, Kim SI, Baek KH, Lee J. Cinnamon bark oil and its components inhibit biofilm formation and toxin production. Int J Food Microbiol. 2015; 195:30-9. [DOI:10.1016/j.ijfoodmicro.2014.11.028] [PMID]
- Fabricant DS, Farnsworth NR. The value of plants used in traditional medicine for drug discovery. Environ Health Perspect. 2001; 109 Suppl 1(Suppl 1):69-75. [DOI:10.1289/ehp.01109s169] [PMID]
- Masuku M, Mozirandi W, Mukanganyama S. Evaluation of the Antibacterial and Antibiofilm Effects of Ethyl Acetate Root Extracts from Vernonia adoensis (Asteraceae) against Pseudomonas aeruginosa. ScientificWorldJournal. 2023; 2023:5782656. [DOI:10.1155/2023/5782656] [PMID]
- Mozirandi W, Mukanganyama S. Antibacterial Activity and Mode of Action of Vernonia adoensis (Asteraceae) Extracts against Staphylococcus aureus and Pseudomonas aeruginosa. J Biol Act Prod Nat. 2017; 7(5):341-57. [DOI:10.1080/22311866.2017.1378922]
- Luo X, Jiang Y, Fronczek FR, Lin C, Izevbigie EB, Lee KS. Isolation and structure determination of a sesquiterpene lactone (vernodalinol) from Vernonia amygdalina extracts. Pharm Biol. 2011; 49(5):464-70. [DOI:10.3109/13880209.2010.523429] [PMID]
- Abay SM, Lucantoni L, Dahiya N, Dori G, Dembo EG, Esposito F, et al. Plasmodium transmission blocking activities of Vernonia amygdalina extracts and isolated compounds. Malar J. 2015; 14:288. [PMID]
- Mozirandi W, Tagwireyi D, Mukanganyama S. Evaluation of antimicrobial activity of chondrillasterol isolated from Vernonia adoensis (Asteraceae). BMC Complement Altern Med. 2019; 19(1):249. [DOI:10.1186/s12906-019-2657-7] [PMID]
- Swamy TA, Obey J, Mutuku NC. Phytochemical analysis of Vernonia adoensis leaves and roots used as a traditional medicinal plants in Kenya. Int J Pharm Biol Sci. 2013; 3(3):46-52. [Link]
- Gull J, Sultana B, Anwar F, Naseer R, Ashraf M, Ashrafuzzaman M. Variation in antioxidant attributes at three ripening stages of guava (Psidium guajava L.) fruit from different geographical regions of Pakistan. Molecules. 2012; 17(3):3165-80. [DOI:10.3390/molecules17033165] [PMID]
- Mphahlele RR, Stander MA, Fawole OA, Opara UL. Effect of fruit maturity and growing location on the postharvest contents of flavonoids, phenolic acids, vitamin C and antioxidant activity of pomegranate juice (cv. Wonderful). Sci Hortic (Amsterdam). 2014; 179:36-45. [DOI:10.1016/j.scienta.2014.09.007]
- Okeke MI, Iroegbu CU, Eze EN, Okoli AS, Esimone CO. Evaluation of extracts of the root of Landolphia owerrience for antibacterial activity. J Ethnopharmacol. 2001; 78(2-3):119-27. [DOI:10.1016/S0378-8741(01)00307-5] [PMID]
- Broschat SL, Call DR, Kuhn EA, Loge FJ. Comparison of the reflectance and crystal violet assays for measurement of bioflm formation by Enterococcus. Biofilms. 2005; 2(3):177-81. [Link]
- Chaieb K, Kouidhi B, Jrah H, Mahdouani K, Bakhrouf A. Antibacterial activity of Thymoquinone, an active principle of Nigella sativa and its potency to prevent bacterial biofilm formation. BMC Complement Altern Med. 2011; 11:29.[DOI:10.1186/1472-6882-11-29] [PMID]
- Ramage G, López-Ribot JL. Techniques for antifungal susceptibility testing of Candida albicans biofilms. Methods Mol Med. 2005; 118:71-9. [DOI:10.1385/1-59259-943-5:071] [PMID]
- Davies DG, Parsek MR, Pearson JP, Iglewski BH, Costerton JW, Greenberg EP. The involvement of cell-to-cell signals in the development of a bacterial biofilm. Science. 1998; 280(5361):295-8. [DOI:10.1126/science.280.5361.295] [PMID]
- Ekow Thomford N, Dzobo K, Adu F, Chirikure S, Wonkam A, Dandara C. Bush mint (Hyptis suaveolens) and spreading hogweed (Boerhavia diffusa) medicinal plant extracts differentially affect activities of CYP1A2, CYP2D6 and CYP3A4 enzymes. J Ethnopharmacol. 2018; 211:58-69. [DOI:10.1016/j.jep.2017.09.023] [PMID]
- Akiyama K, Chikayama E, Yuasa H, Shimada Y, Tohge T, Shinozaki K, et al. PRIMe: A Web site that assembles tools for metabolomics and transcriptomics. In Silico Biol. 2008; 8(3-4):339-45. [PMID]
- Moco S, Bino RJ, Vorst O, Verhoeven HA, de Groot J, van Beek TA, et al. A liquid chromatography-mass spectrometry-based metabolome database for tomato. Plant Physiol. 2006; 141(4):1205-18. [DOI:10.1104/pp.106.078428] [PMID]
- Stoodley P, Sauer K, Davies DG, Costerton JW. Biofilms as complex differentiated communities. Annu Rev Microbiol. 2002; 56:187-209. [DOI:10.1146/annurev.micro.56.012302.160705] [PMID]
- Clatworthy AE, Pierson E, Hung DT. Targeting virulence: A new paradigm for antimicrobial therapy. Nat Chem Biol. 2007; 3(9):541-8. [DOI:10.1038/nchembio.2007.24] [PMID]
- Davies DG, Marques CN. A fatty acid messenger is responsible for inducing dispersion in microbial biofilms. J Bacteriol. 2009; 191(5):1393-403. [DOI:10.1128/JB.01214-08] [PMID]
- Kumar PP, Kumaravel S, Lalitha C. Screening of antioxidant activity, total phenolics and GC-MS study of Vitex negundo. African J Biochem Res. 2009; 4:191-5. [Link]
- Jancy Rani PM, Kannan P, Kumaravel S. Screening of antioxidant activity, total phenolics and gas chromatograph and mass spectrometer (GC-MS) study of Delonix regia. Afr J Biochem Res. 2011; 5(12):341-7. [Link]
- Costa DC, Costa HS, Albuquerque TG, Ramos F, Castilho MC, Sanches-Silva A. Advances in phenolic compounds analysis of aromatic plants and their potential applications. Trends Food Sci Technol. 2015; 45(2):336-54. [DOI:10.1016/j.tifs.2015.06.009]
- Kampkötter A, Gombitang Nkwonkam C, Zurawski RF, Timpel C, Chovolou Y, Wätjen W, et al. Effects of the flavonoids kaempferol and fisetin on thermotolerance, oxidative stress and FoxO transcription factor DAF-16 in the model organism Caenorhabditis elegans. Arch Toxicol. 2007; 81(12):849-58. [DOI:10.1007/s00204-007-0215-4] [PMID]
- Al-Numair KS, Chandramohan G, Veeramani C, Alsaif MA. Ameliorative effect of kaempferol, a flavonoid, on oxidative stress in streptozotocin-induced diabetic rats. Redox Rep. 2015; 20(5):198-209. [DOI:10.1179/1351000214Y.0000000117] [PMID]
- Park MJ, Lee EK, Heo HS, Kim MS, Sung B, Kim MK, et al. The anti-inflammatory effect of kaempferol in aged kidney tissues: the involvement of nuclear factor-kappaB via nuclear factor-inducing kinase/IkappaB kinase and mitogen-activated protein kinase pathways. J Med Food. 2009; 12(2):351-8. [DOI:10.1089/jmf.2008.0006] [PMID]
- Nguyen TT, Tran E, Ong CK, Lee SK, Do PT, Huynh TT, et al. Kaempferol-induced growth inhibition and apoptosis in A549 lung cancer cells is mediated by activation of MEK-MAPK. J Cell Physiol. 2003; 197(1):110-21. [DOI:10.1002/jcp.10340] [PMID]
- Abu-Reidah IM, Ali-Shtayeh MS, Jamous RM, Arraez-Roman D, Segura-Carretero A. Comprehensive metabolite profiling of Arum palaestinum (Araceae) leaves by using liquid chromatography–tandem mass spectrometry. Food Res Int. 2015; 70:74-86. [DOI:10.1016/j.foodres.2015.01.023]
- Ozçelik B, Kartal M, Orhan I. Cytotoxicity, antiviral and antimicrobial activities of alkaloids, flavonoids, and phenolic acids. Pharm Biol. 2011; 49(4):396-402. [DOI:10.3109/13880209.2010.519390] [PMID]
- Hossan S, Rahman S, Bashar ABMA, Jahan R. Rosmarinic acid : A review of its anticancer action. World J Phamarcy Pharm Sci. 2014; 3:57-70. [Link]
- Murtaza G, Sajjad A, Mehmood Z, Shah SH, Siddiqi AR. Possible molecular targets for therapeutic applications of caffeic acid phenethyl ester in inflammation and cancer. J Food Drug Anal. 2015; 23(1):11-18. [DOI:10.1016/j.jfda.2014.06.001] [PMID]
- Husain SR, Cilurd J, Cillard P. Hydroxyl radical scavenging activity of flavonoids. Phytochemistry. 1987; 26(9):2489-91.[DOI:10.1016/S0031-9422(00)83860-1]
- Jnawali HN, Lee E, Jeong KW, Shin A, Heo YS, Kim Y. Anti-inflammatory activity of rhamnetin and a model of its binding to c-Jun NH2-terminal kinase 1 and p38 MAPK. J Nat Prod. 2014; 77(2):258-63. [DOI:10.1021/np400803n] [PMID]
- Dirscherl K, Karlstetter M, Ebert S, Kraus D, Hlawatsch J, Walczak Y, et al. Luteolin triggers global changes in the microglial transcriptome leading to a unique anti-inflammatory and neuroprotective phenotype. J Neuroinflammation. 2010; 7:3. [DOI:10.1186/1742-2094-7-3] [PMID]
- Byun S, Lee KW, Jung SK, Lee EJ, Hwang MK, Lim SH, et al. Luteolin inhibits protein kinase C(epsilon) and c-Src activities and UVB-induced skin cancer. Cancer Res. 2010; 70(6):2415-23. [DOI:10.1158/0008-5472.CAN-09-4093] [PMID]
- Wollenweber E. Flavonoid excretion in betula species. Biochem Physiol Pflanz. 1974; 166(5):425-8. [Link]
- Lou Z, Wang H, Zhu S, Zhang M, Gao Y, Ma C, et al. Improved extraction and identification by ultra performance liquid chromatography tandem mass spectrometry of phenolic compounds in burdock leaves. J Chromatogr A. 2010; 1217(16):2441-6. [DOI:10.1016/j.chroma.2009.12.022] [PMID]
- Liang YZ, Xie P, Chan K. Quality control of herbal medicines. J Chromatogr B Analyt Technol Biomed Life Sci. 2004; 812(1-2):53-70. [DOI:10.1016/j.jchromb.2004.08.041] [PMID]
- Nikam PH, Kareparamban J, Jadhav A, Kadam V. Future trends in standardization of herbal drugs. J Appl Pharm Sci. 2012; 2(6):38-44. [Link]
Type of Study: Original Research |
Subject:
Ehtnopharmacology
| Rights and permissions | |
 |
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License. |

This is an open access article distributed under the terms of the Creative Commons Attribution License (CC-By-NC), which permits use, distribution, and reproduction in any medium, provided the original work is properly cited and is not used for commercial purposes.
Contact Information
