

Volume 11, Issue 4 (2025)
Pharm Biomed Res 2025, 11(4): 347-358 |
Back to browse issues page



Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Mahmoodi M, Mehranjani M S, Bakhshi S. The Impact of N-acetylcysteine Supplementation on Spermatogenic Recovery and Biochemical Markers in Dexamethasone-administered Mice. Pharm Biomed Res 2025; 11 (4) :347-358
URL: http://pbr.mazums.ac.ir/article-1-705-en.html
URL: http://pbr.mazums.ac.ir/article-1-705-en.html


1- Department of Biology, Faculty of Science, Arak University, Arak, Iran.
Full-Text [PDF 1005 kb]
(68 Downloads)
| Abstract (HTML) (182 Views)
Full-Text: (46 Views)
Introduction
Male infertility and its reflection on reproduction have become a concern in recent years [1]. Toxins and medicines can interfere with the reproductive system [2]. Dexamethasone (DEX), a synthetic glucocorticoid, is widely prescribed in inflammatory disorders, autoimmune diseases [3], congenital hyperplasia of the adrenal glands, cerebral edema, shock, rheumatism, detecting Cushing syndrome, endogenous depression, as well as anti-nausea and anti-vomiting agents in patients undergoing chemotherapy [4]. It can suppress the immune system [5, 6], and also affects the hypothalamic-pituitary-gonadal hormone axis, causing significant changes in the level of sex hormones secretion [7, 8]. DEX induces oxidative stress, increases lipid peroxidation and reduces glutathione peroxidase in testicular tissue which ultimately inhibits testosterone secretion; reduce sperm count and motility and leads to a reduction in fertility rate [6, 9, 10]. N-acetylcysteine (NAC) is a derivative of the amino acid cysteine and a glutathione precursor, the major intracellular antioxidant, that decreases during oxidative stress. Therefore NAC consumption, as an antioxidant, can increase the level of glutathione [11], and can be effective in cardiovascular disease, human immunodeficiency virus infections, liver toxicity, and metal poisoning [12]. On the other hand, NAC can reduce apoptosis in male germ cells [13], decrease malondialdehyde (MDA) levels and increase glutamine peroxidation [14, 15]. It also improves sperm motility, semen volume, and viscosity [16]. The protective effects of NAC are mainly attributed to its function as a precursor for glutathione synthesis, which enhances intracellular antioxidant capacity and maintains redox homeostasis by neutralizing reactive oxygen species (ROS). In addition to this indirect effect, NAC directly scavenges free radicals, stabilizes mitochondrial function, and modulates apoptosis-related signalling pathways. By preventing oxidative damage and inhibiting pro-apoptotic cascades, NAC helps preserve testicular structure, supports the function of Leydig and Sertoli cells (SC), and promotes normal steroidogenic processes and spermatogenesis. Considering what was said, we aimed to investigate the influence of NAC, as an antioxidant compound, on toxic effects induced by DEX on spermatogenesis and histopathological alterations of testicular tissue in NMRI mice
Materials and Methods
Animals and treatments
In the present study, 24 adult male NMRI mice weighing 35±2 g were obtained from the Pasteur Institute of Iran and maintained under standard conditions (12 h light/12 h dark cycle, 21±2 °C) in the animal house of Arak University, with free access to food and water. The mice were randomly assigned to 4 study groups (each 6 mice): Control, DEX (IP injections with the dose of 7 mg/kg/d) (Sigma company, Germany), NAC (IP injections with the dose of 100 mg/kg/d) (Sigma company, USA), and DEX+NAC. Treatment lasted for 7 days. The selected doses of DEX [17-19] and NAC [20, 21] were based on previous studies.
Sample collection
Twenty-four hours after the final injection, the animals were weighed and humanely euthanized. The cauda epididymis was carefully excised, placed in 1.5 mL of Ham’s F10 medium, and incubated at 37 °C for 5 min to obtain a sperm suspension for parameter evaluation. For biochemical analyses, blood samples were collected directly from the left ventricle, and their plasma was separated by centrifugation. The left testes were weighed, fixed in modified Davidson’s fluid, and stained using the Heidenhain’s azan method for stereological examination. The right testes were frozen in liquid nitrogen for 10 min and subsequently stored at −80 °C for the assessment of daily sperm production (DSP) and testicular MDA levels.
Evaluation of sperm motility
After pipetting, one drop of the sperm suspension was loaded onto a Neubauer counting chamber. The fields were studied using a light microscope at 200× magnification, and the percentage of motile, progressive motile, and immotile sperms were expressed according to the World Health Organization (WHO) recommendations [22].
Stereological study
The orientator technique was applied to prepare isotropic uniform random (IUR) sections of the testes. In this method, each testis was randomly positioned on a φ clock, which was divided into 9 equal sectors. A number between 1 and 9 was randomly selected, and a cut was made along the chosen line to divide the testis into two parts. The first half was then placed on a θ clock—divided into 9 unequal sectors—with its cut surface aligned along the 0–0 axis. A second random number was selected, and another cut was made parallel to that number. The remaining portion from the second cut was placed perpendicularly on the θ clock so that its cut surface coincided with the 0–0 axis, followed by another parallel incision through a randomly selected number. After standard tissue processing, serial sections with thicknesses of 5 and 20 μm were obtained using a Leitz 1512 microtome and subsequently stained with the Heidenhain’s azan technique [23, 24].
Shrinkage calculation
To determine the extent of testicular shrinkage, 3 randomly selected tissue samples were obtained from the IUR regions using a trocar. For each sample, two vertical diameters were measured, and their average was used to calculate the mean radius, referred to as the pre-fixation radius (rbefore). Following fixation, tissue processing, sectioning, and staining procedures, the mean radius of each sample was measured again and recorded as the post-fixation radius (rafter). The extent of testicular shrinkage for each specimen was then determined using the following equation and by applying it to the initial volume of the testicle (immersion volume), the absolute volume of the testis (Vtotal) was estimated [25] (Equation 1).
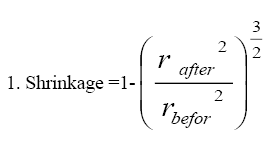
Calculating the number of spermatogenic, sertoli and leydig cells
The optical dissector technique along with an unbiased counting frame was applied to quantify spermatocytes, round and elongated spermatids, spermatogonia, as well as the Leydig and SCs. Random microscopic fields at 100× magnification were selected from tissue sections with a thickness of 20 μm, and a microcator (ND 221 B, Heidenhain, Germany) was utilized for the counting process. The numerical density (Nv) of various cell types was then calculated using the Equation 2.
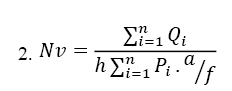
Where ΣQi is the total number of cells counted, h is the tissue thickness that is considered for counting, a/f is the area of the counting frame at the true tissue scale and ΣPi is the total number of points superimposed on the selected fields. Subsequently, to determine the absolute total number of cells (Ntotal), the numerical density (Nv) is multiplied by the total volume (Vtotal) (Equation 3):
3. Ntotal=Nv×Vtotal [25]
Evaluating the spermatogenesis index
Histological assessment was performed on 5-μm tissue sections using an Olympus BX41TE optical microscope (Japan) at 400× magnification. For each animal, all morphometric indices (spermiogenesis (SP), SC, meiosis (M), tubular differentiation indexes [TDI]) were quantified through systematic random sampling of 100 seminiferous tubule cross-sections.
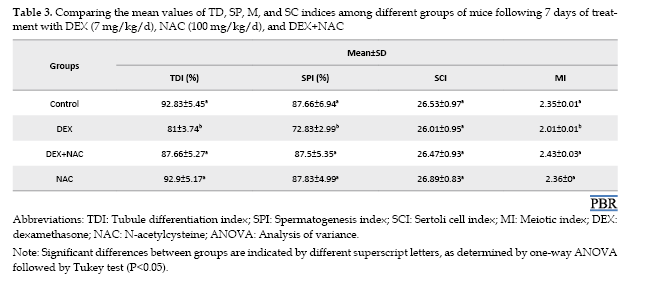
The spermiogenesis index (SPI) represents the percentage of seminiferous tubules that contain mature spermatozoa. The SC index (SCI) describes the ratio between germ cells and SCs. The meiosis index (MI) indicates the proportion of round spermatids relative to primary spermatocytes. Finally, the TDI is defined as the percentage of seminiferous tubules that exhibit at least three different germ cell types derived from type A spermatogonia [18].
Estimating the sperm tail length (STL)
To measure STL, a sperm suspension was first prepared and stained with eosin-nigrosin. A drop of the stained sample was spread onto a glass slide and examined under a light microscope (Olympus BX41TE, Japan) equipped with a digital camera (Olympus DP12, Japan) at 400× magnification. Using a test grid, a total of 200 spermatozoa were evaluated. The STL was then determined through stereological techniques applicable to length estimation in two-dimensional (2D) images. Microscopic fields were selected using a systematic random sampling approach. In accordance with stereological principles, approximately 100–200 probe interactions were recorded, such as counting 100–200 sperm heads within the counting frame or 100–200 intersections of the Merz grid with sperm tails. The mean STL was subsequently calculated using the Equation 4:
4. ΣL (total tails)=(π/2)×(a/l)×(1/asf)×ΣI
L (tail)=ΣL/ΣN
In the Merz grid, the constant “a/l” is determined as follows: The area of each basic tile on the grid is equal to X multiplied by Y. Therefore, the Merz grid constant “a/l” is calculated using the Equation 5:
5. (X×Y)/(π×d)
The term “asf” represents the ratio of the area of a basic tile to the area of the counting frame. “ΣI” denotes the total number of intersections between the sperm tails and the semicircles, while “ΣN” indicates the total number of sperms counted within the unbiased counting frame [24].
DSP
The right testes were first weighed and decapsulated, after which the testicular parenchyma was homogenized in 2 mL of NaCl solution containing 0.05% Triton X-100. To visualize the spermatids, a few drops of 1% eosin were added to the homogenate. Subsequently, 5 μL aliquot of the suspension was transferred to a Neubauer chamber, and the sperm cells were examined under a light microscope at 400× magnification.
The DSP was calculated according to the Equation 6:
6. X= a×5×b×10000,
where “X” represents the number of spermatids per milliliter of sample, “a” is the count of spermatids in 5 squares, 5 is a constant, and “b” denotes the dilution factor.
The testicular spermatid number (TSN) was obtained using the Equation 7:
7. (TSN)= X/W
where W is the testicular weight (in g).
Finally, the DSP was determined as Equation 8:
8. DSP= TSN/4.84
where 4.84 corresponds to the average number of days required for developing spermatids to progress through the 14–16 cell stage of spermatogenesis in mice [18, 26, 27].
Biochemical studies
Evaluating Serum MDA
The serum MDA level was measured using the method described by Buege and Aust. For this purpose, a TCA–TBA–HCl reagent was prepared, consisting of 15% (g/mL) trichloroacetic acid (TCA), 0.375% (g/mL) thiobarbituric acid (TBA), and 25% normal hydrochloric acid (HCl). One part of the serum sample was mixed with two parts of this reagent, and the mixture was incubated in a water bath at 100 °C for 15 min. Following incubation, the samples were rapidly cooled in cold water and centrifuged for 10 min. The supernatant was then collected, and its absorbance was read at 532 nm using a spectrophotometer (T80C, PG Instruments Ltd, London, UK). The MDA concentration was determined from the absorbance values using an extinction coefficient of 1.56×10⁵ M⁻¹ cm⁻¹ and expressed as nmol/mL [28, 29].
Testicular MDA assay
The right testis was weighed and homogenized with KCl solution for 2 min. Then, it was mixed with a TBA-TCA solution. The reaction mixture was incubated in a water bath at 100 °C for 15 min, then rapidly cooled and centrifuged at 10000 g for 10 min. The absorbance of the resulting supernatant was measured at 532 nm using a spectrophotometer. The MDA content in the testicular tissue homogenate was quantified and expressed as nmol/mg of tissue [15].
Testosterone assay
Testosterone levels were measured using an enzyme-linked immunosorbent assay (ELISA) with the Monobind Testosterone ELISA kit (EIA-37K5J5, Monobind Inc., Lake Forest, California, USA), following the protocol provided by the manufacturer.
Statistical analysis
Data analysis was performed using SPSS software, version 16. One-way analysis of variance (ANOVA) followed by Tukey post hoc test was used to compare group means, with differences considered statistically significant at P<0.05.
Results
Histopathological findings
Histological studies of the testis tissue in the control group showed a normal testis tissue and the seminiferous tubules with normal spermatogenesis (Figure 1A). In the DEX group, decreased spermatogenesis and increased inter-tissue with edema were found. The testicular tissue of this group demonstrated marked degenerative changes, such as decreased germinal epithelial height, vacuolization, epithelial rupture in the seminiferous tubules, and reduced sperm accumulation within the tubular lumen (Figure 1B). Microscopic evaluation of the testes in the DEX+NAC group revealed that the majority of histopathological alterations induced by DEX were ameliorated, with the seminiferous tubules showing an almost normal epithelial configuration (Figure 1C). The NAC group demonstrated tubules structurally similar to those of the control group (Figure 1D).
.PNG)
Body and testis weight
Statistical analysis showed no significant differences in the mean body weights betrween the control, NAC, DEX, and DEX+NAC groups after the treatment period (P>0.05). Also, the mean testis weight did not differ significantly between the experimental groups (P>0.05) (Table 1).
.PNG)
The mean number of spermatocytes, round and long spermatids, spermatogonia, leydig and SCs
In the DEX-treated group, the mean numbers of spermatocytes, round spermatids, elongated spermatids, and Leydig cells were significantly reduced compared with the other experimental groups (P<0.01). These parameters increased significantly in the DEX+NAC group to the control level (P>0.05). No significant differences were observed in the mean numbers of spermatogonia and SCs among the groups (P>0.05) (Table 2).
.PNG)
Spermatogenesis indices
The M (P<0.05), SP (P<0.05), and TD (P<0.003) indices showed a significant reduction in the DEX-treated group compared with the control group. In contrast, these parameters were markedly improved in the DEX+NAC group, reaching values comparable to those of the control. No significant difference was detected in the SCI among the four groups (P>0.05) (Table 3).
Sperm motility
A significant decline in the mean percentage of progressively motile sperm (P<0.001) and a concomitant rise in non-progressive and immotile sperm (P<0.001) were recorded in the DEX-treated group compared with the control. Co-administration of NAC with DEX significantly ameliorated these changes, resulting in values comparable to the control level (P>0.05) (Table 4).
.PNG)
STL
Comparing the mean STL revealed no significant difference between the DEX group and the control, NAC, and DEX+NAC groups (P>0.05) (Table 4).
DSP rate
A significant decline in the mean DSP was observed in the DEX group relative to the control group (P<0.01). In the DEX+NAC group, this parameter increased significantly, reaching values comparable to those of the control group (P>0.05) (Table 4).
Lipid peroxidation level
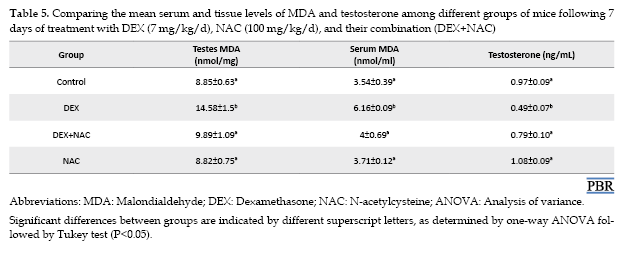
A significant increase in serum and testicular MDA concentrations was observed in the DEX group relative to the control group (P<0.001). Co-administration of NAC with DEX significantly reduced MDA levels, restoring them to control values (P>0.05) (Table 5).
Testosterone hormone
The testosterone level showed a significant decline in the DEX-treated group compared with the control (P<0.05), whereas co-administration of NAC with DEX significantly increased testosterone levels, reaching values not significantly different from those of the control (P>0.05) (Table 5).
Discussion
Our findings revealed that DEX treatment in adult mice significantly decreased the mean number of spermatocytes, round and long spermatids, and Leydig cells, along with reductions in the indices of M, SP and TD, which is consistent with findings reported in previous studies [18, 30, 31]. DEX appears to exert its deleterious effects on testicular function through the activation of the Fas/FasL signalling pathway, stimulation of caspase activity, and upregulation of pro-apoptotic proteins such as Bax. Furthermore, DEX can induce mitochondrial membrane depolarization and increase the production of ROS, leading to oxidative stress. These alterations ultimately contribute to impaired steroidogenesis and a reduction in testosterone levels [10, 31–34].
It has been shown that high doses of DEX can result in apoptosis of SCs [17, 35]. However, in the present study, the mean number of spermatogonia, SCs and SCI in both DEX and control groups showed no significant differences, which is in accordance with the results obtained by Jorge and et al. [17]. This result could be due to the low-dose of DEX used in the current research or the short-term treatment or even the higher weight of mice studied in our case.
The DEX group exhibited a significantly lower proportion of progressively motile spermatozoa than the control group. Notably, the pronounced elevation in the proportions of non-progressive and immotile spermatozoa was detected in the DEX-treated mice relative to controls, which aligns with the results of previous research [36, 37]. DEX induces membrane lipid peroxidation through the production of free radicals [38], leading to a loss of membrane fluidity, reduced activity of membrane-associated enzymes and ion channels, receptor inactivation, and nonspecific ion permeability, which can explain the gradual loss of sperm motility [39]. ROS production not only reduces the phosphorylation of axonal proteins but also damages the mitochondrial membrane resulting in the ATP depletion, which is the source of energy for sperm motility [39] ultimately leading to poor sperm motility [40]. Despite the reduction in sperm motility, our data showed no significant difference in the STL between the DEX and control groups, which is in accordance with the findings of our previous study [18].
A significant decline in the DSP rate was observed in mice treated with DEX relative to the control group, which has been reported by our previous study [18]. Generally, DSP depends on the number of somatic cells, including Sertoli and Leydig cells, and also the number of spermatogenic cells [41]. The Leydig cells are also responsible for providing the blood testosterone levels [42], which is crucial for the development and differentiation of sperm cells during spermatogenesis; Therefore, it can be inferred that DEX disrupts the process of spermatogenesis and reduces DSP by inducing apoptosis in spermatogenic, Sertoli, and Leydig cells [19]. Accumulating evidence along with the results of this study have also shown that DEX can lead to a decrease in the blood testosterone levels [10, 18, 43, 44] not only by disturbing the hypothalamus and the pituitary axis, but also by directly affecting the testis Leydig cells via transcriptional inhibition of genes involved in the enzymatic pathways of testosterone synthesis [44, 45], which could be considered another cause for the reduction found in the DSP in this study.
The findings of this study also demonstrate a significant increase in serum and testicular MDA levels in the DEX-treated group relative to controls, presumably resulting from oxidative stress [38] and increase of lipid peroxidation, these findings are in line with the previous researches [17, 18, 46].
Based on the results, no significant differences were observed in the mean testicular or body weights between the groups at the end of the treatment period. Some studies confirm our results [17, 47]; however, others have reported a reduction in testis [18] and body weight [48] in DEX-treated mice. These different outcomes might be related to the stage of treatment which could be either before or after puberty, the duration of treatment, mice strain, and the consumed dose of DEX.
NAC increases the levels of intracellular glutathione and antioxidant enzymes [49] and reduces ROS production [50] which contributes significantly to safeguarding cellular structures against oxidative injury and lipid peroxidation [51]. It can also suppress the expression of pro-apoptotic genes [14] by inhibiting the activities of c-Jun N-terminal kinase, MAP kinase p38, SAPK/INK, c-fos pathway, and NF-κB, thereby preventing cell death [52]. All this together can prevent testis tissue damage, apoptosis and degeneration of germ and Leydig cells [53], which explains the obtained results in this study where NAC significantly increased the number of germ, Sertoli and Leydig cells, spermatogenesis indices, DSP, sperm motility, testosterone levels and also decreased the serum and testis level of MDA to the control level in mice treated with DEX.
Conclusion
In the present study, NAC ameliorated the detrimental effects of DEX on spermatogenesis indices, sperm characteristics and DSP due to its antioxidant properties and its ability in increasing the levels of glutathione and testosterone. Therefore, the consumption of NAC in therapeutic regimens containing DEX is suggested.
Ethical Considerations
Compliance with ethical guidelines
This study was approved by the Ethics Committee of Arak University of Medical Sciences, Arak, Iran (Code: IR.ARAKMU.1397.091).
Funding
This research did not receive any grant from funding agencies in the public, commercial, or non-profit sectors.
Authors' contributions
Data analysis: Monireh Mahmoodi and Malek Soleimani Mehranjani; Writing, review and editing: Monireh Mahmoodi; Manuscript revision: Malek Soleimani Mehranjani; Methodology, data collection and validation, drafting the manuscript, and laboratory experiments: Sepideh Bakhshi.
Conflict of interest
The authors declared no conflict of interest.
Acknowledgments
The authors gratefully acknowledge the cooperation and support of Arak University in conducting this research.
References
Male infertility and its reflection on reproduction have become a concern in recent years [1]. Toxins and medicines can interfere with the reproductive system [2]. Dexamethasone (DEX), a synthetic glucocorticoid, is widely prescribed in inflammatory disorders, autoimmune diseases [3], congenital hyperplasia of the adrenal glands, cerebral edema, shock, rheumatism, detecting Cushing syndrome, endogenous depression, as well as anti-nausea and anti-vomiting agents in patients undergoing chemotherapy [4]. It can suppress the immune system [5, 6], and also affects the hypothalamic-pituitary-gonadal hormone axis, causing significant changes in the level of sex hormones secretion [7, 8]. DEX induces oxidative stress, increases lipid peroxidation and reduces glutathione peroxidase in testicular tissue which ultimately inhibits testosterone secretion; reduce sperm count and motility and leads to a reduction in fertility rate [6, 9, 10]. N-acetylcysteine (NAC) is a derivative of the amino acid cysteine and a glutathione precursor, the major intracellular antioxidant, that decreases during oxidative stress. Therefore NAC consumption, as an antioxidant, can increase the level of glutathione [11], and can be effective in cardiovascular disease, human immunodeficiency virus infections, liver toxicity, and metal poisoning [12]. On the other hand, NAC can reduce apoptosis in male germ cells [13], decrease malondialdehyde (MDA) levels and increase glutamine peroxidation [14, 15]. It also improves sperm motility, semen volume, and viscosity [16]. The protective effects of NAC are mainly attributed to its function as a precursor for glutathione synthesis, which enhances intracellular antioxidant capacity and maintains redox homeostasis by neutralizing reactive oxygen species (ROS). In addition to this indirect effect, NAC directly scavenges free radicals, stabilizes mitochondrial function, and modulates apoptosis-related signalling pathways. By preventing oxidative damage and inhibiting pro-apoptotic cascades, NAC helps preserve testicular structure, supports the function of Leydig and Sertoli cells (SC), and promotes normal steroidogenic processes and spermatogenesis. Considering what was said, we aimed to investigate the influence of NAC, as an antioxidant compound, on toxic effects induced by DEX on spermatogenesis and histopathological alterations of testicular tissue in NMRI mice
Materials and Methods
Animals and treatments
In the present study, 24 adult male NMRI mice weighing 35±2 g were obtained from the Pasteur Institute of Iran and maintained under standard conditions (12 h light/12 h dark cycle, 21±2 °C) in the animal house of Arak University, with free access to food and water. The mice were randomly assigned to 4 study groups (each 6 mice): Control, DEX (IP injections with the dose of 7 mg/kg/d) (Sigma company, Germany), NAC (IP injections with the dose of 100 mg/kg/d) (Sigma company, USA), and DEX+NAC. Treatment lasted for 7 days. The selected doses of DEX [17-19] and NAC [20, 21] were based on previous studies.
Sample collection
Twenty-four hours after the final injection, the animals were weighed and humanely euthanized. The cauda epididymis was carefully excised, placed in 1.5 mL of Ham’s F10 medium, and incubated at 37 °C for 5 min to obtain a sperm suspension for parameter evaluation. For biochemical analyses, blood samples were collected directly from the left ventricle, and their plasma was separated by centrifugation. The left testes were weighed, fixed in modified Davidson’s fluid, and stained using the Heidenhain’s azan method for stereological examination. The right testes were frozen in liquid nitrogen for 10 min and subsequently stored at −80 °C for the assessment of daily sperm production (DSP) and testicular MDA levels.
Evaluation of sperm motility
After pipetting, one drop of the sperm suspension was loaded onto a Neubauer counting chamber. The fields were studied using a light microscope at 200× magnification, and the percentage of motile, progressive motile, and immotile sperms were expressed according to the World Health Organization (WHO) recommendations [22].
Stereological study
The orientator technique was applied to prepare isotropic uniform random (IUR) sections of the testes. In this method, each testis was randomly positioned on a φ clock, which was divided into 9 equal sectors. A number between 1 and 9 was randomly selected, and a cut was made along the chosen line to divide the testis into two parts. The first half was then placed on a θ clock—divided into 9 unequal sectors—with its cut surface aligned along the 0–0 axis. A second random number was selected, and another cut was made parallel to that number. The remaining portion from the second cut was placed perpendicularly on the θ clock so that its cut surface coincided with the 0–0 axis, followed by another parallel incision through a randomly selected number. After standard tissue processing, serial sections with thicknesses of 5 and 20 μm were obtained using a Leitz 1512 microtome and subsequently stained with the Heidenhain’s azan technique [23, 24].
Shrinkage calculation
To determine the extent of testicular shrinkage, 3 randomly selected tissue samples were obtained from the IUR regions using a trocar. For each sample, two vertical diameters were measured, and their average was used to calculate the mean radius, referred to as the pre-fixation radius (rbefore). Following fixation, tissue processing, sectioning, and staining procedures, the mean radius of each sample was measured again and recorded as the post-fixation radius (rafter). The extent of testicular shrinkage for each specimen was then determined using the following equation and by applying it to the initial volume of the testicle (immersion volume), the absolute volume of the testis (Vtotal) was estimated [25] (Equation 1).
Calculating the number of spermatogenic, sertoli and leydig cells
The optical dissector technique along with an unbiased counting frame was applied to quantify spermatocytes, round and elongated spermatids, spermatogonia, as well as the Leydig and SCs. Random microscopic fields at 100× magnification were selected from tissue sections with a thickness of 20 μm, and a microcator (ND 221 B, Heidenhain, Germany) was utilized for the counting process. The numerical density (Nv) of various cell types was then calculated using the Equation 2.
Where ΣQi is the total number of cells counted, h is the tissue thickness that is considered for counting, a/f is the area of the counting frame at the true tissue scale and ΣPi is the total number of points superimposed on the selected fields. Subsequently, to determine the absolute total number of cells (Ntotal), the numerical density (Nv) is multiplied by the total volume (Vtotal) (Equation 3):
3. Ntotal=Nv×Vtotal [25]
Evaluating the spermatogenesis index
Histological assessment was performed on 5-μm tissue sections using an Olympus BX41TE optical microscope (Japan) at 400× magnification. For each animal, all morphometric indices (spermiogenesis (SP), SC, meiosis (M), tubular differentiation indexes [TDI]) were quantified through systematic random sampling of 100 seminiferous tubule cross-sections.
The spermiogenesis index (SPI) represents the percentage of seminiferous tubules that contain mature spermatozoa. The SC index (SCI) describes the ratio between germ cells and SCs. The meiosis index (MI) indicates the proportion of round spermatids relative to primary spermatocytes. Finally, the TDI is defined as the percentage of seminiferous tubules that exhibit at least three different germ cell types derived from type A spermatogonia [18].
Estimating the sperm tail length (STL)
To measure STL, a sperm suspension was first prepared and stained with eosin-nigrosin. A drop of the stained sample was spread onto a glass slide and examined under a light microscope (Olympus BX41TE, Japan) equipped with a digital camera (Olympus DP12, Japan) at 400× magnification. Using a test grid, a total of 200 spermatozoa were evaluated. The STL was then determined through stereological techniques applicable to length estimation in two-dimensional (2D) images. Microscopic fields were selected using a systematic random sampling approach. In accordance with stereological principles, approximately 100–200 probe interactions were recorded, such as counting 100–200 sperm heads within the counting frame or 100–200 intersections of the Merz grid with sperm tails. The mean STL was subsequently calculated using the Equation 4:
4. ΣL (total tails)=(π/2)×(a/l)×(1/asf)×ΣI
L (tail)=ΣL/ΣN
In the Merz grid, the constant “a/l” is determined as follows: The area of each basic tile on the grid is equal to X multiplied by Y. Therefore, the Merz grid constant “a/l” is calculated using the Equation 5:
5. (X×Y)/(π×d)
The term “asf” represents the ratio of the area of a basic tile to the area of the counting frame. “ΣI” denotes the total number of intersections between the sperm tails and the semicircles, while “ΣN” indicates the total number of sperms counted within the unbiased counting frame [24].
DSP
The right testes were first weighed and decapsulated, after which the testicular parenchyma was homogenized in 2 mL of NaCl solution containing 0.05% Triton X-100. To visualize the spermatids, a few drops of 1% eosin were added to the homogenate. Subsequently, 5 μL aliquot of the suspension was transferred to a Neubauer chamber, and the sperm cells were examined under a light microscope at 400× magnification.
The DSP was calculated according to the Equation 6:
6. X= a×5×b×10000,
where “X” represents the number of spermatids per milliliter of sample, “a” is the count of spermatids in 5 squares, 5 is a constant, and “b” denotes the dilution factor.
The testicular spermatid number (TSN) was obtained using the Equation 7:
7. (TSN)= X/W
where W is the testicular weight (in g).
Finally, the DSP was determined as Equation 8:
8. DSP= TSN/4.84
where 4.84 corresponds to the average number of days required for developing spermatids to progress through the 14–16 cell stage of spermatogenesis in mice [18, 26, 27].
Biochemical studies
Evaluating Serum MDA
The serum MDA level was measured using the method described by Buege and Aust. For this purpose, a TCA–TBA–HCl reagent was prepared, consisting of 15% (g/mL) trichloroacetic acid (TCA), 0.375% (g/mL) thiobarbituric acid (TBA), and 25% normal hydrochloric acid (HCl). One part of the serum sample was mixed with two parts of this reagent, and the mixture was incubated in a water bath at 100 °C for 15 min. Following incubation, the samples were rapidly cooled in cold water and centrifuged for 10 min. The supernatant was then collected, and its absorbance was read at 532 nm using a spectrophotometer (T80C, PG Instruments Ltd, London, UK). The MDA concentration was determined from the absorbance values using an extinction coefficient of 1.56×10⁵ M⁻¹ cm⁻¹ and expressed as nmol/mL [28, 29].
Testicular MDA assay
The right testis was weighed and homogenized with KCl solution for 2 min. Then, it was mixed with a TBA-TCA solution. The reaction mixture was incubated in a water bath at 100 °C for 15 min, then rapidly cooled and centrifuged at 10000 g for 10 min. The absorbance of the resulting supernatant was measured at 532 nm using a spectrophotometer. The MDA content in the testicular tissue homogenate was quantified and expressed as nmol/mg of tissue [15].
Testosterone assay
Testosterone levels were measured using an enzyme-linked immunosorbent assay (ELISA) with the Monobind Testosterone ELISA kit (EIA-37K5J5, Monobind Inc., Lake Forest, California, USA), following the protocol provided by the manufacturer.
Statistical analysis
Data analysis was performed using SPSS software, version 16. One-way analysis of variance (ANOVA) followed by Tukey post hoc test was used to compare group means, with differences considered statistically significant at P<0.05.
Results
Histopathological findings
Histological studies of the testis tissue in the control group showed a normal testis tissue and the seminiferous tubules with normal spermatogenesis (Figure 1A). In the DEX group, decreased spermatogenesis and increased inter-tissue with edema were found. The testicular tissue of this group demonstrated marked degenerative changes, such as decreased germinal epithelial height, vacuolization, epithelial rupture in the seminiferous tubules, and reduced sperm accumulation within the tubular lumen (Figure 1B). Microscopic evaluation of the testes in the DEX+NAC group revealed that the majority of histopathological alterations induced by DEX were ameliorated, with the seminiferous tubules showing an almost normal epithelial configuration (Figure 1C). The NAC group demonstrated tubules structurally similar to those of the control group (Figure 1D).
Body and testis weight
Statistical analysis showed no significant differences in the mean body weights betrween the control, NAC, DEX, and DEX+NAC groups after the treatment period (P>0.05). Also, the mean testis weight did not differ significantly between the experimental groups (P>0.05) (Table 1).
The mean number of spermatocytes, round and long spermatids, spermatogonia, leydig and SCs
In the DEX-treated group, the mean numbers of spermatocytes, round spermatids, elongated spermatids, and Leydig cells were significantly reduced compared with the other experimental groups (P<0.01). These parameters increased significantly in the DEX+NAC group to the control level (P>0.05). No significant differences were observed in the mean numbers of spermatogonia and SCs among the groups (P>0.05) (Table 2).
Spermatogenesis indices
The M (P<0.05), SP (P<0.05), and TD (P<0.003) indices showed a significant reduction in the DEX-treated group compared with the control group. In contrast, these parameters were markedly improved in the DEX+NAC group, reaching values comparable to those of the control. No significant difference was detected in the SCI among the four groups (P>0.05) (Table 3).
Sperm motility
A significant decline in the mean percentage of progressively motile sperm (P<0.001) and a concomitant rise in non-progressive and immotile sperm (P<0.001) were recorded in the DEX-treated group compared with the control. Co-administration of NAC with DEX significantly ameliorated these changes, resulting in values comparable to the control level (P>0.05) (Table 4).
STL
Comparing the mean STL revealed no significant difference between the DEX group and the control, NAC, and DEX+NAC groups (P>0.05) (Table 4).
DSP rate
A significant decline in the mean DSP was observed in the DEX group relative to the control group (P<0.01). In the DEX+NAC group, this parameter increased significantly, reaching values comparable to those of the control group (P>0.05) (Table 4).
Lipid peroxidation level
A significant increase in serum and testicular MDA concentrations was observed in the DEX group relative to the control group (P<0.001). Co-administration of NAC with DEX significantly reduced MDA levels, restoring them to control values (P>0.05) (Table 5).
Testosterone hormone
The testosterone level showed a significant decline in the DEX-treated group compared with the control (P<0.05), whereas co-administration of NAC with DEX significantly increased testosterone levels, reaching values not significantly different from those of the control (P>0.05) (Table 5).
Discussion
Our findings revealed that DEX treatment in adult mice significantly decreased the mean number of spermatocytes, round and long spermatids, and Leydig cells, along with reductions in the indices of M, SP and TD, which is consistent with findings reported in previous studies [18, 30, 31]. DEX appears to exert its deleterious effects on testicular function through the activation of the Fas/FasL signalling pathway, stimulation of caspase activity, and upregulation of pro-apoptotic proteins such as Bax. Furthermore, DEX can induce mitochondrial membrane depolarization and increase the production of ROS, leading to oxidative stress. These alterations ultimately contribute to impaired steroidogenesis and a reduction in testosterone levels [10, 31–34].
It has been shown that high doses of DEX can result in apoptosis of SCs [17, 35]. However, in the present study, the mean number of spermatogonia, SCs and SCI in both DEX and control groups showed no significant differences, which is in accordance with the results obtained by Jorge and et al. [17]. This result could be due to the low-dose of DEX used in the current research or the short-term treatment or even the higher weight of mice studied in our case.
The DEX group exhibited a significantly lower proportion of progressively motile spermatozoa than the control group. Notably, the pronounced elevation in the proportions of non-progressive and immotile spermatozoa was detected in the DEX-treated mice relative to controls, which aligns with the results of previous research [36, 37]. DEX induces membrane lipid peroxidation through the production of free radicals [38], leading to a loss of membrane fluidity, reduced activity of membrane-associated enzymes and ion channels, receptor inactivation, and nonspecific ion permeability, which can explain the gradual loss of sperm motility [39]. ROS production not only reduces the phosphorylation of axonal proteins but also damages the mitochondrial membrane resulting in the ATP depletion, which is the source of energy for sperm motility [39] ultimately leading to poor sperm motility [40]. Despite the reduction in sperm motility, our data showed no significant difference in the STL between the DEX and control groups, which is in accordance with the findings of our previous study [18].
A significant decline in the DSP rate was observed in mice treated with DEX relative to the control group, which has been reported by our previous study [18]. Generally, DSP depends on the number of somatic cells, including Sertoli and Leydig cells, and also the number of spermatogenic cells [41]. The Leydig cells are also responsible for providing the blood testosterone levels [42], which is crucial for the development and differentiation of sperm cells during spermatogenesis; Therefore, it can be inferred that DEX disrupts the process of spermatogenesis and reduces DSP by inducing apoptosis in spermatogenic, Sertoli, and Leydig cells [19]. Accumulating evidence along with the results of this study have also shown that DEX can lead to a decrease in the blood testosterone levels [10, 18, 43, 44] not only by disturbing the hypothalamus and the pituitary axis, but also by directly affecting the testis Leydig cells via transcriptional inhibition of genes involved in the enzymatic pathways of testosterone synthesis [44, 45], which could be considered another cause for the reduction found in the DSP in this study.
The findings of this study also demonstrate a significant increase in serum and testicular MDA levels in the DEX-treated group relative to controls, presumably resulting from oxidative stress [38] and increase of lipid peroxidation, these findings are in line with the previous researches [17, 18, 46].
Based on the results, no significant differences were observed in the mean testicular or body weights between the groups at the end of the treatment period. Some studies confirm our results [17, 47]; however, others have reported a reduction in testis [18] and body weight [48] in DEX-treated mice. These different outcomes might be related to the stage of treatment which could be either before or after puberty, the duration of treatment, mice strain, and the consumed dose of DEX.
NAC increases the levels of intracellular glutathione and antioxidant enzymes [49] and reduces ROS production [50] which contributes significantly to safeguarding cellular structures against oxidative injury and lipid peroxidation [51]. It can also suppress the expression of pro-apoptotic genes [14] by inhibiting the activities of c-Jun N-terminal kinase, MAP kinase p38, SAPK/INK, c-fos pathway, and NF-κB, thereby preventing cell death [52]. All this together can prevent testis tissue damage, apoptosis and degeneration of germ and Leydig cells [53], which explains the obtained results in this study where NAC significantly increased the number of germ, Sertoli and Leydig cells, spermatogenesis indices, DSP, sperm motility, testosterone levels and also decreased the serum and testis level of MDA to the control level in mice treated with DEX.
Conclusion
In the present study, NAC ameliorated the detrimental effects of DEX on spermatogenesis indices, sperm characteristics and DSP due to its antioxidant properties and its ability in increasing the levels of glutathione and testosterone. Therefore, the consumption of NAC in therapeutic regimens containing DEX is suggested.
Ethical Considerations
Compliance with ethical guidelines
This study was approved by the Ethics Committee of Arak University of Medical Sciences, Arak, Iran (Code: IR.ARAKMU.1397.091).
Funding
This research did not receive any grant from funding agencies in the public, commercial, or non-profit sectors.
Authors' contributions
Data analysis: Monireh Mahmoodi and Malek Soleimani Mehranjani; Writing, review and editing: Monireh Mahmoodi; Manuscript revision: Malek Soleimani Mehranjani; Methodology, data collection and validation, drafting the manuscript, and laboratory experiments: Sepideh Bakhshi.
Conflict of interest
The authors declared no conflict of interest.
Acknowledgments
The authors gratefully acknowledge the cooperation and support of Arak University in conducting this research.
References
- Kaltsas A, Zachariou A, Dimitriadis F, Chrisofos M, Sofikitis N. Empirical treatments for male infertility: A focus on lifestyle modifications and medicines. Diseases. 2024; 12(9):209. [DOI:10.3390/diseases12090209] [PMID]
- Krzastek SC, Farhi J, Gray M, Smith RP. Impact of environmental toxin exposure on male fertility potential. Transl Androl Urol. 2020; 9(6):2797-813. [DOI:10.21037/tau-20-685] [PMID]
- Dubashynskaya NV, Bokatyi AN, Skorik YA. Dexamethasone conjugates: Synthetic approaches and medical prospects. Biomedicines. 2021; 9(4):341. [DOI:10.3390/biomedicines9040341] [PMID]
- Madamsetty VS, Mohammadinejad R, Uzieliene I, Nabavi N, Dehshahri A, García-Couce J, et al. Dexamethasone: Insights into pharmacological aspects, therapeutic mechanisms, and delivery systems. ACS Biomater Sci Eng. 2022; 8(5):1763-90. [DOI:10.1021/acsbiomaterials.2c00026] [PMID]
- Sinner B. Perioperatives Dexamethason [Perioperative dexamethasone (German)]. Anaesthesist. 2019; 68(10):676-82. [DOI:10.1007/s00101-019-00672-x] [PMID]
- Iorgulescu JB, Gokhale PC, Speranza MC, Eschle BK, Poitras MJ, Wilkens MK, et al. Concurrent dexamethasone limits the clinical benefit of immune checkpoint blockade in glioblastoma. Clin Cancer Res. 2021; 27(1):276-87. [DOI:10.1158/1078-0432.CCR-20-2291] [PMID]
- Ma Y, Fan X, Han J, Cheng Y, Zhao J, Fang W, et al. Critical illness and sex hormones: Response and impact of the hypothalamic-pituitary-gonadal axis. Ther Adv Endocrinol Metab. 2025; 16:20420188251328192.[DOI:10.1177/20420188251328192] [PMID]
- Ignatiuk V, Izvolskaia M, Sharova V, Zakharova L. Disruptions in hypothalamic-pituitary-gonadal axis development and their igg modulation after prenatal systemic inflammation in male rats. Int J Mol Sci. 2023; 24(3):2726. [DOI:10.3390/ijms24032726] [PMID]
- Kong Z, Zhu L, Liu Y, Liu Y, Chen G, Wang H. Effects of different stages, dosages and courses of prenatal dexamethasone exposure on testicular development in mice. Food Chem Toxicol. 2025; 201:115468. [DOI:10.1016/j.fct.2025.115468] [PMID]
- Annie L, Gurusubramanian G, Kumar Roy V. Dexamethasone mediated downregulation of PGC-1α and visfatin regulates testosterone synthesis and antioxidant system in mouse testis. Acta Histochemica. 2019; 121(2):182-8. [DOI:10.1016/j.acthis.2018.12.004] [PMID]
- Tenório MCDS, Graciliano NG, Moura FA, Oliveira ACM, Goulart MOF. N-Acetylcysteine (NAC): Impacts on human health. Antioxidants (Basel). 2021; 10(6):967. [DOI:10.3390/antiox10060967] [PMID]
- Mokra D, Porvaznik I, Mokry J. N-Acetylcysteine in the Treatment of Acute Lung Injury: Perspectives and Limitations. Int J Mol Sci. 2025; 26(6):2657. [DOI:10.3390/ijms26062657] [PMID]
- Abedini Bajgiran F, Khazaei Koohpar Z, Salehzadeh A. Effects of N-Acetylcysteine supplementation on oxidative stress and expression of apoptosis-related genes in testicular tissue of rats exposed to lead. Biol Trace Elem Res. 2023; 201(5):2407-15. [DOI:10.1007/s12011-022-03325-0] [PMID]
- Mohammadi-Sardoo M, Mandegary A, Nabiuni M, Nematollahi-Mahani SN, Amirheidari B. Mancozeb induces testicular dysfunction through oxidative stress and apoptosis: Protective role of N-acetylcysteine antioxidant. Toxicol Ind Health. 2018; 34(11):798-811. [DOI:10.1177/0748233718778397] [PMID]
- Malmir M, Soleimani Mehranjani M, Naderi Noreini S, Faraji T. Protective antioxidant effects of N‐acetylcysteine against impairment of spermatogenesis caused by paranonylphenol. Andrologia. 2018; 50(10):e13114. [DOI:10.1111/and.13114] [PMID]
- Wang Y, Xu Y, Yang L, Yang Y, Guo AL, Han XJ, et al. N-acetylcysteine alleviated tris (2-chloroisopropyl) phosphate-induced sperm motility decline and functional dysfunction in mice through reversing oxidative stress and DNA damage. Ecotoxicol Environ Saf. 2024; 271:116000. [DOI:10.1016/j.ecoenv.2024.116000] [PMID]
- Soleimani Mehranjani M, Azizi M, Sadeghzadeh F. The effect of melatonin on testis histological changes and spermatogenesis indexes in mice following treatment with dexamethasone. Drug Chem Toxicol. 2022; 45(3):1140-9. [DOI:10.1080/01480545.2020.1809672] [PMID]
- Sadeghzadeh F, Mehranjani M, Mahmoodi M. Vitamin C ameliorates the adverse effects of dexamethasone on sperm motility, testosterone level, and spermatogenesis indexes in mice. Hum Exp Toxicol. 2019; 38(4):409-18. [DOI:10.1177/0960327118816137] [PMID]
- Mohamadpour M, Mollajani R, Sarabandi F, Hosseini F, Mohsenkia M, Erfanizadeh M, et al. Protective effect of grape seed extract on dexamethasone-induced testicular toxicity in mice. Crescent J Med Biol Sci. 2020; 7(1):59-65. [Link]
- Turkmen R, Akosman MS, Demirel HH. Protective effect of N-acetylcysteine on MK-801-induced testicular oxidative stress in mice. Biomed Pharmacother. 2019; 109:1988-93. [DOI:10.1016/j.biopha.2018.09.139] [PMID]
- Shittu SA, Shittu ST, Akindele OO, Kunle-Alabi OT, Raji Y. Protective action of N-acetylcysteine on sperm quality in cyclophosphamide-induced testicular toxicity in male Wistar rats. JBRA Assist Reprod. 2019; 23(2):83-90. [DOI:10.5935/1518-0557.20180079] [PMID]
- Björndahl L, Kirkman Brown J; other Editorial Board Members of the WHO Laboratory Manual for the Examination and Processing of Human Semen. The sixth edition of the WHO laboratory manual for the examination and processing of human semen: ensuring quality and standardization in basic examination of human ejaculates. Fertil Steril. 2022; 117(2):246-51. [DOI:10.1016/j.fertnstert.2021.12.012] [PMID]
- Blutke A, Wanke R. Sampling strategies and processing of biobank tissue samples from porcine biomedical models. J Vis Exp. 2018; (133):57276. [DOI:10.3791/57276] [PMID]
- Dehghani F, Sotoude N, Bordbar H, Panjeshahin MR, Karbalay-Doust S. The use of platelet-rich plasma (PRP) to improve structural impairment of rat testis induced by busulfan. Platelets. 2019; 30(4):513-20. [DOI:10.1080/09537104.2018.1478400] [PMID]
- Zamani A, Saki F, Hatami N, Koohpeyma F. Stereological assessment of the effects of vitamin D deficiency on the rat testis. BMC Endocr Disord. 2020; 20(1):162. [DOI:10.1186/s12902-020-00642-0] [PMID]
- Siervo GEML, Mariani NAP, Silva AAS, Punhagui-Umbelino APF, Costa IRD, Andrade AD, et al. Low dose of cyclosporine A disrupts sperm parameters and testosterone levels reversibly in mice. Toxicol Appl Pharmacol. 2023; 460:116374. [DOI:10.1016/j.taap.2023.116374] [PMID]
- Tootian Z, Fazelipour S, Goodarzi N, Arab HA. The effect of pure phenol on sperm parameters and fertility rate in male mice. Iran J Vet Med. 2015; 9(4):295-301. [Link]
- Buege JA, Aust SD. [30] Microsomal lipid peroxidation. In: Methods in enzymology. Amsterdam: Elsevier; 1978. [DOI:10.1016/S0076-6879(78)52032-6]
- Tutar E, Pesen B, Coşkun ZM, Bolkent S. Oxidative stress status in testis of type-2 diabetic rats treated with Delta-9-tetrahydrocannabinol. Eur J Biol. 2021; 80(2):91-6. [Link]
- Abouzaripour M, Daneshi E, Miri S. Effect of Nigella sativa on dexamethasone-induced testicular toxicity in mice. Avicenna J Phytomed. 2024; 14(5):520-6. [DOI:10.22038/AJP.2023.23584] [PMID]
- Chaimontri C, Arun S, Kamollerd T, Lapyuneyong N, Taoto C, Innoi S, et al. Low sperm quality and increased testicular caspases in chronic stress mice induced by dexamethasone. Braz J Biol. 2025; 85:e289850. [DOI:10.1590/1519-6984.289850] [PMID]
- Hajimirza SP, Forouzesh F, Shabani M. [Effect of dexamethasone on Fas/FasL and Bax/Bcl2 mRNA expression in human colorectal Cancer HT-29 cell line (Persian)]. Res Med. 2020; 44(2):352-9. [Link]
- Deng S, Dai G, Chen S, Nie Z, Zhou J, Fang H, et al. Dexamethasone induces osteoblast apoptosis through ROS-PI3K/AKT/GSK3β signaling pathway. Biomed Pharmacother. 2019; 110:602-8. [DOI:10.1016/j.biopha.2018.11.103] [PMID]
- Alahmar AT, Al Jothery AH, Al-Daami QJ, Abbas A, Al-Hassnawi AT. The effect of coenzyme Q10 on dexamethasone-induced oxidative stress in rats testes. Med J Babylon. 2023; 20(1):130-5. [Link]
- Ren L, Zhang Y, Xin Y, Chen G, Sun X, Chen Y, et al. Dysfunction in sertoli cells participates in glucocorticoid-induced impairment of spermatogenesis. Mol Reprod Dev. 2021; 88(6):405-15. [DOI:10.1002/mrd.23515] [PMID]
- Mahmoodi M, Soleimani Mehranjani M, Kheradmand N. [Protective effect of pentoxifylline on sperm and biochemical parameters against dexamethasone-induced toxicity in mice (Persian)]. J Mazandaran Univ Med Sci. 2020; 30(185):113-24.[Link]
- Soleimani Mehranjani M, Mohammadi S M. [The protective effect of quercetin on sperm parameters and serum biochemical factors in adult mice treated with dexamethasone (Persian)]. Qom Univ Med Sci J. 2019; 13(2):27-38. [DOI:10.29252/qums.13.2.27]
- Nassan MA, Soliman MM, Aldhahrani A, El‐Saway HB, Swelum AA. Ameliorative impacts of Allium cepa Linnaeus aqueous extract against testicular damage induced by dexamethasone. Andrologia. 2021; 53(4):e13955. [DOI:10.1111/and.13955] [PMID]
- Gallo A, Esposito MC, Tosti E, Boni R. Sperm motility, oxidative status, and mitochondrial activity: Exploring correlation in different species. Antioxidants (Basel). 2021; 10(7):1131. [DOI:10.3390/antiox10071131] [PMID]
- Ribeiro JC, Nogueira-Ferreira R, Amado F, Alves MG, Ferreira R, Oliveira PF. Exploring the role of oxidative stress in sperm motility: A proteomic network approach. Antioxid Redox Signal. 2022; 37(7-9):501-20. [DOI:10.1089/ars.2021.0241] [PMID]
- Lara NL, Santos IC, Costa GM, Cordeiro-Junior DA, Almeida AC, Madureira AP, et al. Duration of spermatogenesis and daily sperm production in the rodent Proechimys guyannensis. Zygote. 2016; 24(5):783-93. [DOI:10.1017/S0967199416000137] [PMID]
- Zirkin BR, Papadopoulos V. Leydig cells: formation, function, and regulation. Biol Reprod. 2018; 99(1):101-11. [DOI:10.1093/biolre/ioy059] [PMID]
- Zhang J, Hu G, Huang B, Zhuo D, Xu Y, Li H, Zhan X, Ge RS, Xu Y. Dexamethasone suppresses the differentiation of stem Leydig cells in rats in vitro. BMC Pharmacol Toxicol. 2019; 20(1):32. [DOI:10.1186/s40360-019-0312-z] [PMID]
- Hasona NA. Grape seed extract attenuates dexamethasone-induced testicular and thyroid dysfunction in male albino rats. Andrologia. 2018; 50(5):e13002. [DOI:10.1111/and.13002] [PMID]
- Liu M, Chen B, Pei L, Zhang Q, Zou Y, Xiao H, et al Decreased H3K9ac level of StAR mediated testicular dysplasia induced by prenatal dexamethasone exposure in male offspring rats. Toxicology. 2018; 408:1-10. [PMID]
- Dare JB, Ojewale AO, Olaniyan OT, Adole JA, Okotie GE, Akintayo CO, et al. Adansonia digitata aqueous leaf extract ameliorates dexamethasone-induced testicular injury in male Wistar rats. Asian Pac J Reprod. 2021; 10(3):113-20. [Link]
- Azimi Zangabad E, Shomali T, Roshangar L. Effects of pharmacological doses of niacin on subacute glucocorticoid‐induced testicular damage in rats. Pharmacol Res Perspect. 2023; 11(5):e01128. [PMID]
- Filippopoulou F, Habeos GI, Rinotas V, Sophocleous A, Sykiotis GP, Douni E, et al. Dexamethasone administration in mice leads to less body weight gain over time, lower serum glucose, and higher insulin levels independently of NRF2. Antioxidants (Basel). 2021; 11(1):4. [DOI:10.3390/antiox11010004] [PMID]
- Schmitt B, Vicenzi M, Garrel C, Denis FM. Effects of N-acetylcysteine, oral glutathione (GSH) and a novel sublingual form of GSH on oxidative stress markers: A comparative crossover study. Redox Biol. 2015; 6:198-205. [DOI:10.1016/j.redox.2015.07.012] [PMID]
- Xie C, Yi J, Lu J, Nie M, Huang M, Rong J, et al. N‐acetylcysteine reduces ROS‐mediated oxidative DNA damage and PI3K/Akt pathway activation induced by Helicobacter pylori infection. Oxid Med Cell Longev. 2018; 2018:1874985. [DOI:10.1155/2018/1874985] [PMID]
- Malmir M, Faraji T, Noreini N, Mehranjani M. Protective antioxidant effects of N-acetylcysteine on testicular tissue and serum testosterone in paranonylphenol-treated mice (a stereological analysis). Reprod Syst Sex Disord. 2018; 7(2):225. [Link]
- Ghafarizadeh A, Malmir M, Naderi Noreini S, Faraji T. Antioxidant effects of N-acetylcysteine on the male reproductive system: A systematic review. Andrologia. 2021; 53(1):e13898. [DOI:10.1111/and.13898] [PMID]
- Shahrzad E, Shariati M, Naimi S, Edalatmanesh MA. Protective effect of N-acetylcysteine on changes in serum levels of Pituitary–Gonadal axis hormones and testicular tissue in acrylamide-treated adult rats. Adv Hum Biol. 2020; 10(1):16-21. [DOI:10.4103/AIHB.AIHB_65_19]
Type of Study: Original Research |
Subject:
Toxicology
| Rights and permissions | |
 |
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License. |

This is an open access article distributed under the terms of the Creative Commons Attribution License (CC-By-NC), which permits use, distribution, and reproduction in any medium, provided the original work is properly cited and is not used for commercial purposes.
Contact Information
